

Mitotic degradation of human thymidine kinase 1 is dependent on the anaphase-promoting complex/cyclosome-CDH1-mediated pathway
- PMID: 14701726
- PMCID: PMC343798
- DOI: 10.1128/MCB.24.2.514-526.2004
Mitotic degradation of human thymidine kinase 1 is dependent on the anaphase-promoting complex/cyclosome-CDH1-mediated pathway
Abstract
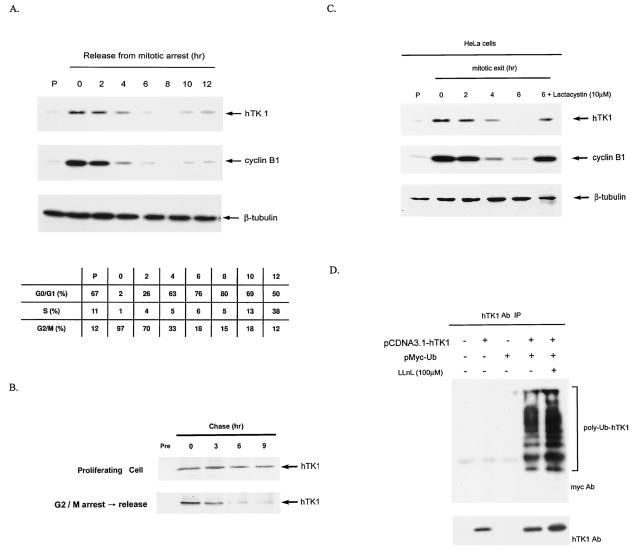
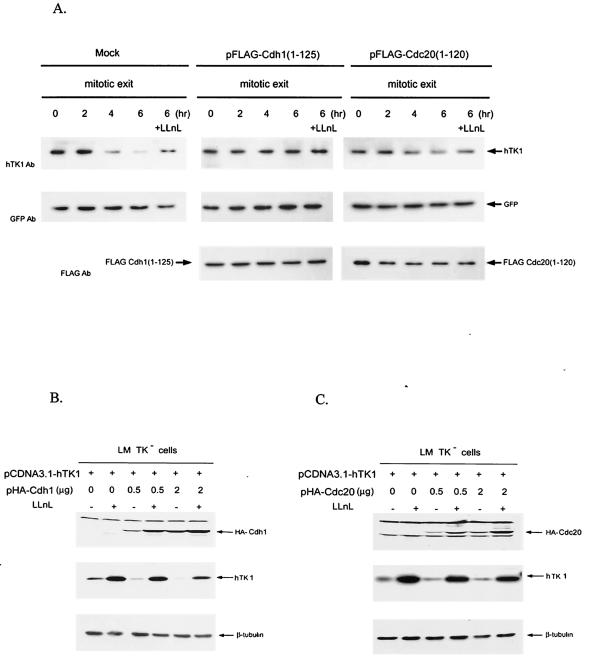
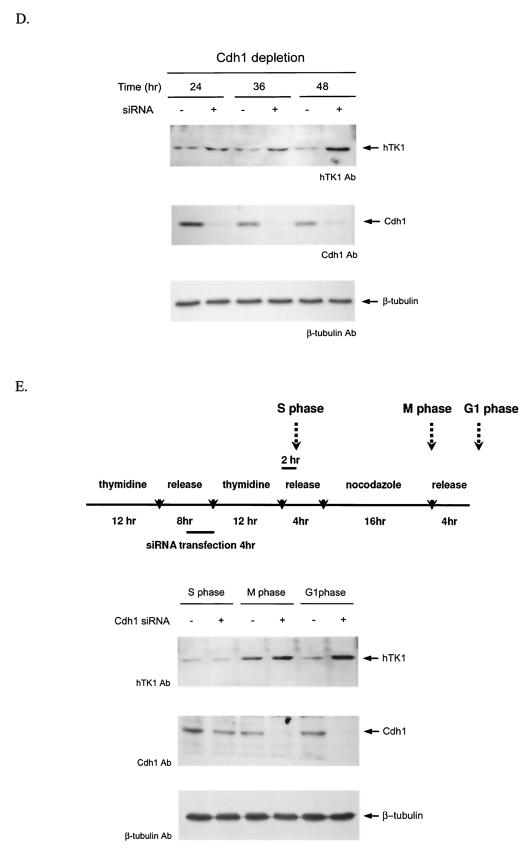
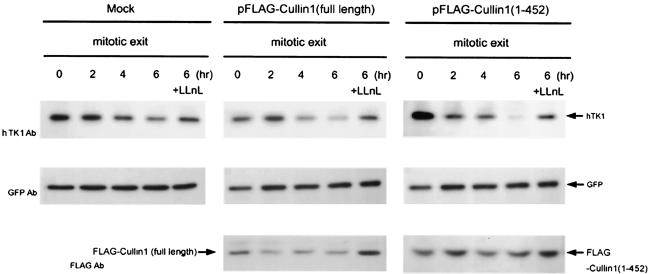
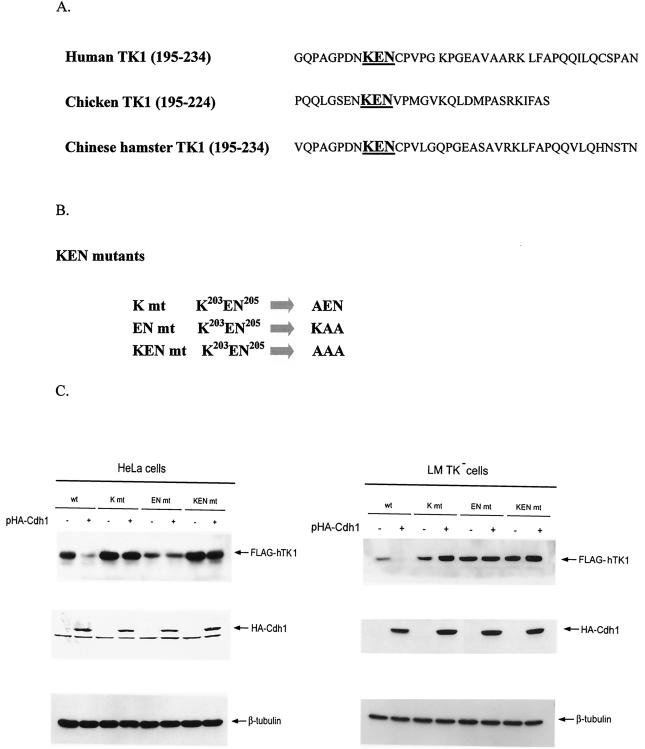
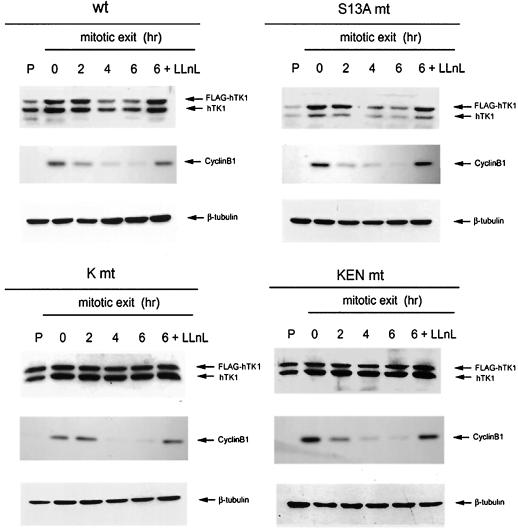
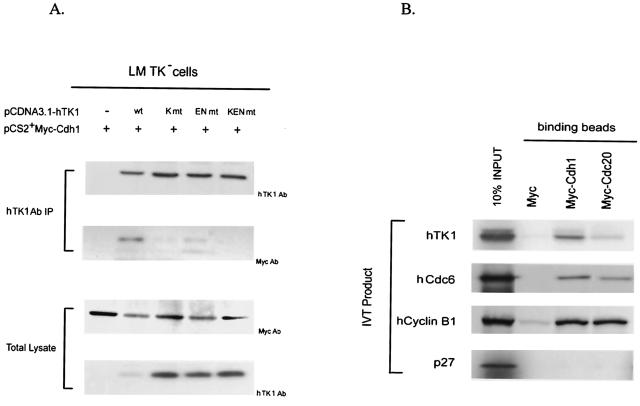
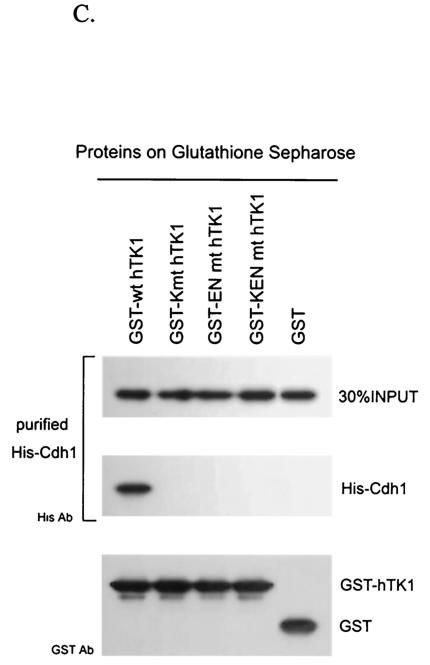
The expression of human thymidine kinase 1 (hTK1) is highly dependent on the growth states and cell cycle stages in mammalian cells. The amount of hTK1 is significantly increased in the cells during progression to the S and M phases, and becomes barely detectable in the early G(1) phase by a proteolytic control during mitotic exit. This tight regulation is important for providing the correct pool of dTTP for DNA synthesis at the right time in the cell cycle. Here, we investigated the mechanism responsible for mitotic degradation of hTK1. We show that hTK1 is degraded via a ubiquitin-proteasome pathway in mammalian cells and that anaphase-promoting complex/cyclosome (APC/C) activator Cdh1 is not only a necessary but also a rate-limiting factor for mitotic degradation of hTK1. Furthermore, a KEN box sequence located in the C-terminal region of hTK1 is required for its mitotic degradation and interaction capability with Cdh1. By in vitro ubiquitinylation assays, we demonstrated that hTK1 is targeted for degradation by the APC/C-Cdh1 ubiquitin ligase dependent on this KEN box motif. Taken together, we concluded that activation of the APC/C-Cdh1 complex during mitotic exit controls timing of hTK1 destruction, thus effectively minimizing dTTP formation from the salvage pathway in the early G(1) phase of the cell cycle in mammalian cells.
Figures
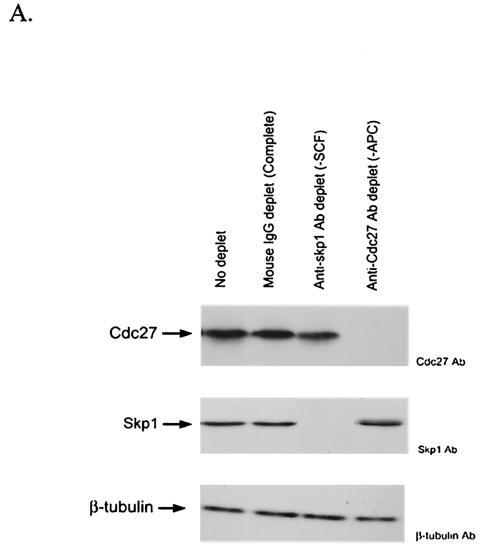
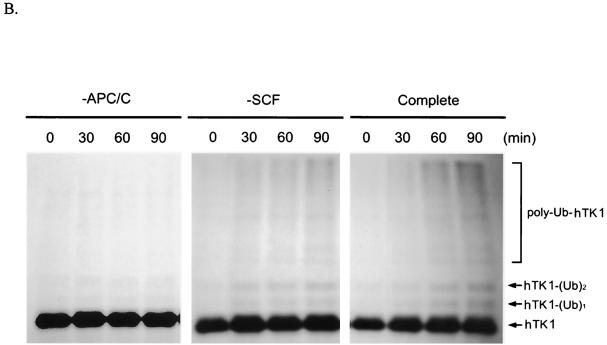
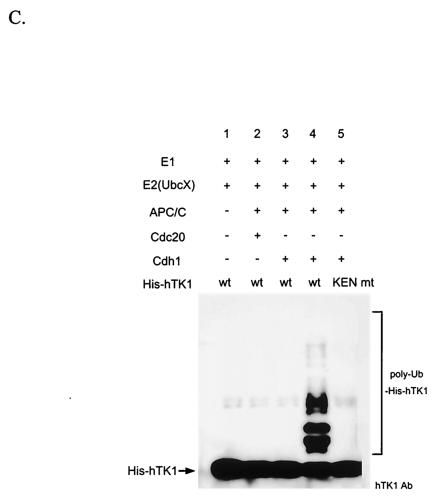
References
-
- Bello, L. J. 1974. Regulation of thymidine kinase synthesis in human cells. Exp. Cell Res. 89:263-274. - PubMed
-
- Chang, Z. F., D. Y. Huang, and N. C. Hsue. 1994. Differential phosphorylation of human thymidine kinase in proliferating and M phase-arrested human cells. J. Biol. Chem. 269:21249-21254. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous