

Nuclear reorganization of mammalian DNA synthesis prior to cell cycle exit
- PMID: 14701733
- PMCID: PMC343811
- DOI: 10.1128/MCB.24.2.595-607.2004
Nuclear reorganization of mammalian DNA synthesis prior to cell cycle exit
Abstract
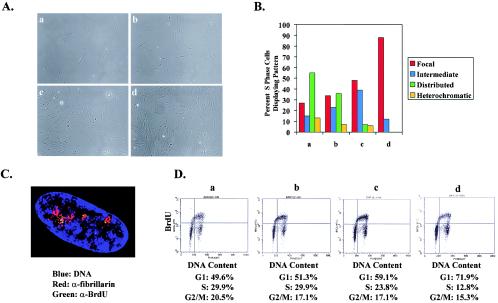
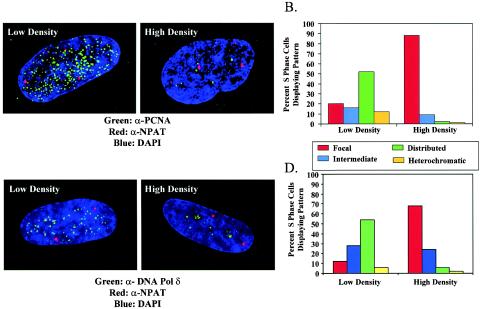
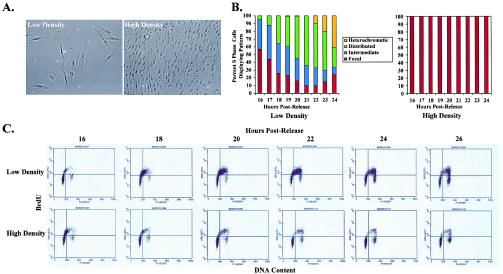
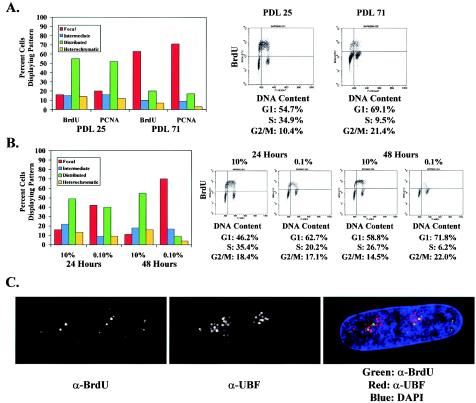
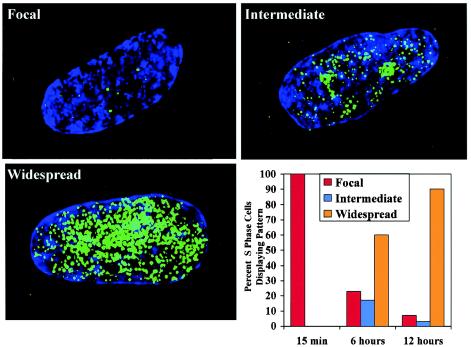
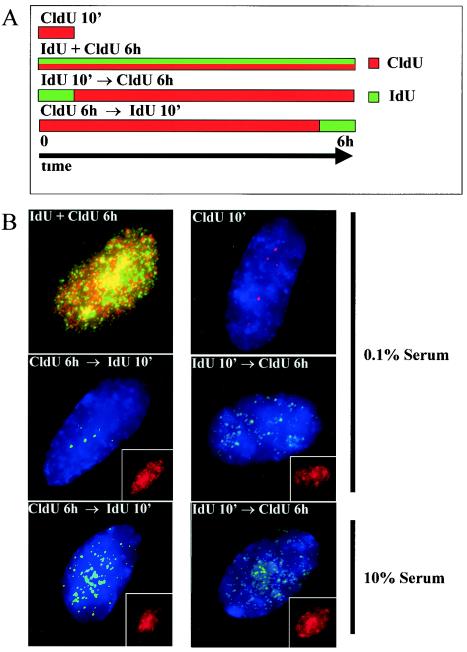
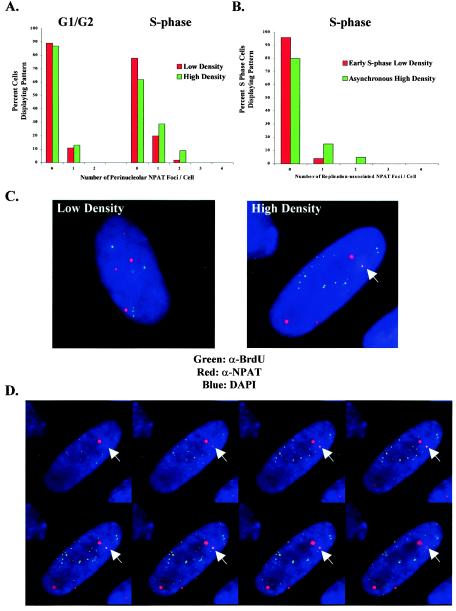
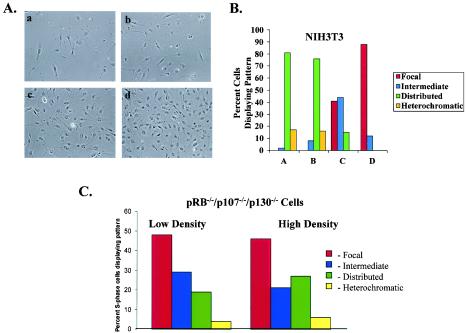
In primary mammalian cells, DNA replication initiates in a small number of perinucleolar, lamin A/C-associated foci. During S-phase progression in proliferating cells, replication foci distribute to hundreds of sites throughout the nucleus. In contrast, we find that the limited perinucleolar replication sites persist throughout S phase as cells prepare to exit the cell cycle in response to contact inhibition, serum starvation, or replicative senescence. Proteins known to be involved in DNA synthesis, such as PCNA and DNA polymerase delta, are concentrated in perinucleolar foci throughout S phase under these conditions. Moreover, chromosomal loci are redirected toward the nucleolus and overlap with the perinucleolar replication foci in cells poised to undergo cell cycle exit. These same loci remain in the periphery of the nucleus during replication under highly proliferative conditions. These results suggest that mammalian cells undergo a large-scale reorganization of chromatin during the rounds of DNA replication that precede cell cycle exit.
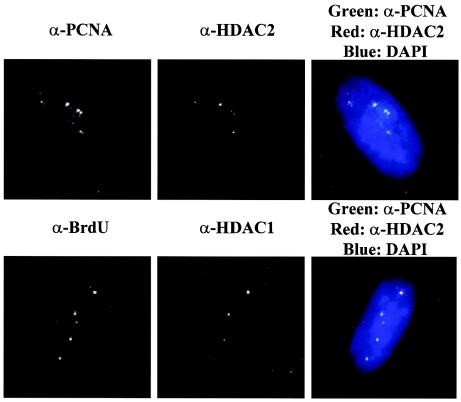
Figures
References
-
- Almouzni, G., and A. P. Wolffe. 1993. Replication-coupled chromatin assembly is required for the repression of basal transcription in vivo. Genes Dev. 7:2033-2047. - PubMed
-
- Bannister, A. J., P. Zegerman, J. F. Partridge, E. A. Miska, J. O. Thomas, R. C. Allshire, and T. Kouzarides. 2001. Selective recognition of methylated lysine 9 on histone H3 by the HP1 chromo domain. Nature 410:120-124. - PubMed
-
- Berezney, R., D. D. Dubey, and J. A. Huberman. 2000. Heterogeneity of eukaryotic replicons, replicon clusters, and replication foci. Chromosoma 108:471-484. - PubMed
-
- Chuang, L. S.-H., H. I. Ian, T. W. Koh, H. H. Ng, G. Xu, and B. F. Li. 1997. Human DNA-(cytosine-5) methyltransferase-PCNA complex as a target for p21WAF1. Science 277:1996-2000. - PubMed
-
- Dimitrova, D. S., and R. Berezney. 2002. The spatio-temporal organization of DNA replication sites is identical in primary, immortalized, and transformed mammalian cells. J. Cell Sci. 115:4037-4051. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous