

Role of the C-terminal domain of RNA polymerase II in U2 snRNA transcription and 3' processing
- PMID: 14701755
- PMCID: PMC343789
- DOI: 10.1128/MCB.24.2.846-855.2004
Role of the C-terminal domain of RNA polymerase II in U2 snRNA transcription and 3' processing
Abstract
U small nuclear RNAs (snRNAs) and mRNAs are both transcribed by RNA polymerase II (Pol II), but the snRNAs have unusual TATA-less promoters and are neither spliced nor polyadenylated; instead, 3' processing is directed by a highly conserved 3' end formation signal that requires initiation from an snRNA promoter. Here we show that the C-terminal domain (CTD) of Pol II is required for efficient U2 snRNA transcription, as it is for mRNA transcription. However, CTD kinase inhibitors, such as 5,6-dichloro-1-beta-D-ribofuranosylbenzimidazole (DRB) and 1-(5-isoquinolinesulfonyl)-2-methylpiperazine (H7), that block mRNA elongation do not affect U2 transcription, although 3' processing of the U2 primary transcript is impaired. We show further that U2 transcription is preferentially inhibited by low doses of UV irradiation or actinomycin D, which induce CTD kinase activity, and that UV inhibition can be rescued by treatment with DRB or H7. We propose that Pol II complexes transcribing snRNAs and mRNAs have distinct CTD phosphorylation patterns. mRNA promoters recruit factors including kinases that hyperphosphorylate the CTD, and the CTD in turn recruits proteins needed for mRNA splicing and polyadenylation. We predict that snRNA promoters recruit factors including a CTD kinase(s) whose snRNA-specific phosphorylation pattern recruits factors required for promoter-coupled 3' end formation.
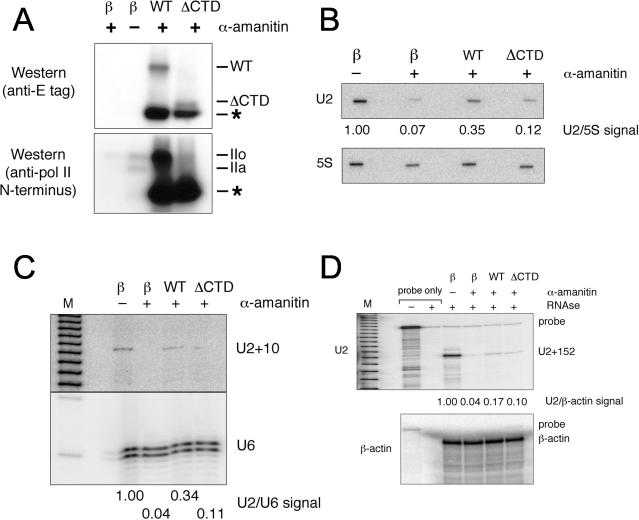
Figures





References
-
- Allison, L. A., M. Moyle, M. Shales, and C. J. Ingles. 1985. Extensive homology among the largest subunits of eukaryotic and prokaryotic RNA polymerases. Cell 42:599-610. - PubMed
-
- Baskaran, R., G. G. Chiang, T. Mysliwiec, G. D. Kruh, and J. Y. Wang. 1997. Tyrosine phosphorylation of RNA polymerase II carboxyl-terminal domain by the Abl-related gene product. J. Biol. Chem. 272:18905-18909. - PubMed
-
- Bender, K., C. Blattner, A. Knebel, M. Iordanov, P. Herrlich, and H. J. Rahmsdorf. 1997. UV-induced signal transduction. J. Photochem. Photobiol. B 37:1-17. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases