

A form of long-lasting, learning-related synaptic plasticity in the hippocampus induced by heterosynaptic low-frequency pairing
- PMID: 14711997
- PMCID: PMC321771
- DOI: 10.1073/pnas.2237201100
A form of long-lasting, learning-related synaptic plasticity in the hippocampus induced by heterosynaptic low-frequency pairing
Abstract
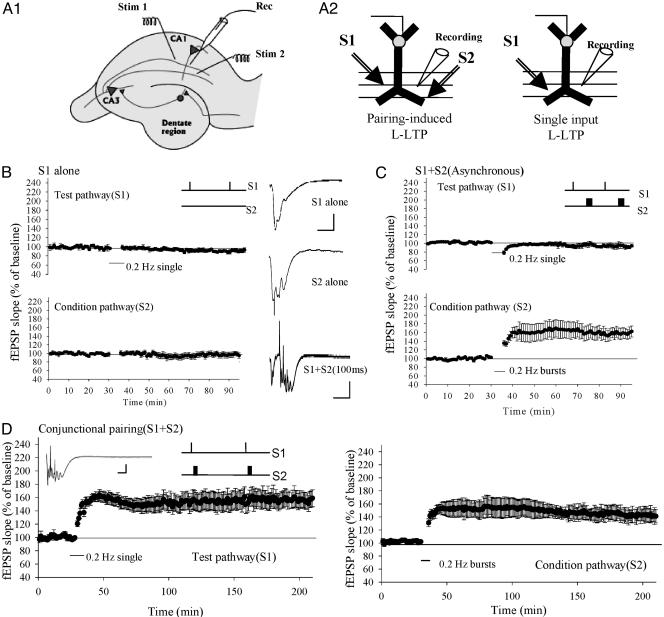
The late, transcription- and translation-dependent phase of long-term synaptic potentiation (L-LTP) at the Schaffer collateral synapse of the hippocampus is an experimental model of the synaptic plasticity underlying long-lasting memory formation. L-LTP is typically induced by homosynaptic tetanic stimulation; but associative forms of learning are likely to require the heterosynaptic pairing of stimuli. Here we describe L-LTP elicited by such heterosynaptic pairing at the Schaffer collateral synapse in mice. We find that repeated stimulation of one pathway at low frequency (0.2 Hz), which does not by itself induce synaptic potentiation, will produce long-lasting synaptic plasticity when paired with a brief conditioning burst applied to an independent afferent pathway. The induction of heterosynaptic L-LTP is associative and critically depends on the precise time interval of pairing: simultaneous, conjunctional pairing induces L-LTP; in contrast, delayed pairing induces short-lasting early-phase LTP. Heterosynaptically induced early-phase LTP could be depotentiated by repeatedly presenting unpaired test stimuli, whereas L-LTP could not. This heterosynaptically induced L-LTP requires PKA and protein synthesis. In addition, heterosynaptically induced L-LTP is impaired in transgenic mice that express KCREB (a dominant negative inhibitor of adenosine 3'5'-cyclic monophosphate response element-binding protein-mediated transcription) in the hippocampus. These mice have previously been shown to be impaired in spatial memory but have normal L-LTP as induced by a conventional homosynaptic tetanic protocol. These data suggest that at least in some instances this L-LTP-inducing protocol may better model behaviorally relevant information storage and the in vivo mechanisms underlying long-lasting memories.
Figures



Similar articles
-
Homosynaptic and heterosynaptic inhibition of synaptic tagging and capture of long-term potentiation by previous synaptic activity.J Neurosci. 2005 Aug 3;25(31):7221-31. doi: 10.1523/JNEUROSCI.0909-05.2005. J Neurosci. 2005. PMID: 16079404 Free PMC article.
-
Brief theta-burst stimulation induces a transcription-dependent late phase of LTP requiring cAMP in area CA1 of the mouse hippocampus.Learn Mem. 1997 Jul-Aug;4(2):230-43. doi: 10.1101/lm.4.2.230. Learn Mem. 1997. PMID: 10456066
-
Translational control of hippocampal synaptic plasticity and memory by the eIF2alpha kinase GCN2.Nature. 2005 Aug 25;436(7054):1166-73. doi: 10.1038/nature03897. Nature. 2005. PMID: 16121183 Free PMC article.
-
LTD, LTP, and the sliding threshold for long-term synaptic plasticity.Hippocampus. 1996;6(1):35-42. doi: 10.1002/(SICI)1098-1063(1996)6:1<35::AID-HIPO7>3.0.CO;2-6. Hippocampus. 1996. PMID: 8878740 Review.
-
Regulation of late-phase LTP and long-term memory in normal and aging hippocampus: role of secreted proteins tPA and BDNF.Ageing Res Rev. 2004 Nov;3(4):407-30. doi: 10.1016/j.arr.2004.07.002. Ageing Res Rev. 2004. PMID: 15541709 Review.
Cited by
-
Combination effects of sodium butyrate and pyridoxine treatment on cell proliferation and neuroblast differentiation in the dentate gyrus of D-galactose-induced aging model mice.Neurochem Res. 2012 Jan;37(1):223-31. doi: 10.1007/s11064-011-0597-9. Epub 2011 Oct 5. Neurochem Res. 2012. PMID: 21984169
-
Ketogenic diet and BHB rescue the fall of long-term potentiation in an Alzheimer's mouse model and stimulates synaptic plasticity pathway enzymes.Commun Biol. 2024 Feb 16;7(1):195. doi: 10.1038/s42003-024-05860-z. Commun Biol. 2024. PMID: 38366025 Free PMC article.
-
Acute Ethanol Exposure Enhances Synaptic Plasticity in the Dorsal Striatum in Adult Male and Female Rats.Brain Plast. 2020 Dec 29;6(1):113-122. doi: 10.3233/BPL-190097. Brain Plast. 2020. PMID: 33680850 Free PMC article.
-
Effectiveness assessment of repetitive transcranial alternating current stimulation with concurrent EEG and fNIRS measurement.Health Inf Sci Syst. 2023 Aug 2;11(1):35. doi: 10.1007/s13755-023-00233-y. eCollection 2023 Dec. Health Inf Sci Syst. 2023. PMID: 37545487 Free PMC article.
-
Impaired bidirectional synaptic plasticity and procedural memory formation in striatum-specific cAMP response element-binding protein-deficient mice.J Neurosci. 2006 Mar 8;26(10):2808-13. doi: 10.1523/JNEUROSCI.5406-05.2006. J Neurosci. 2006. PMID: 16525060 Free PMC article.
References
-
- Martin, S. J., Grimwood, P. D. & Morris, R. G. (2000) Annu. Rev. Neurosci. 23, 649–711. - PubMed
-
- Kang, H, Welcher, A. A., Shelton, D. & Schuman, E. M. (1997) Neuron 19, 653–664. - PubMed
-
- Patterson, S. L., Pittenger, C., Morozov, A., Martin, K. C., Scanlin, H., Drake, C. & Kandel, E. R. (2001) Neuron 32, 123–140. - PubMed
-
- Pittenger, C., Huang, Y.-Y., Paletzki, R. F., Bourtchouladze, R., Scanlin, H., Vronskaya, S. & Kandel, E. R. (2002) Neuron 34, 447–462. - PubMed
-
- Huang, Y.-Y., Nguyen, P. V., Abel, T. & Kandel E. R. (1996) Learn. Mem. 3, 74–85. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
