

Coregulator-dependent facilitation of chromatin occupancy by GATA-1
- PMID: 14715908
- PMCID: PMC327128
- DOI: 10.1073/pnas.0307612100
Coregulator-dependent facilitation of chromatin occupancy by GATA-1
Abstract
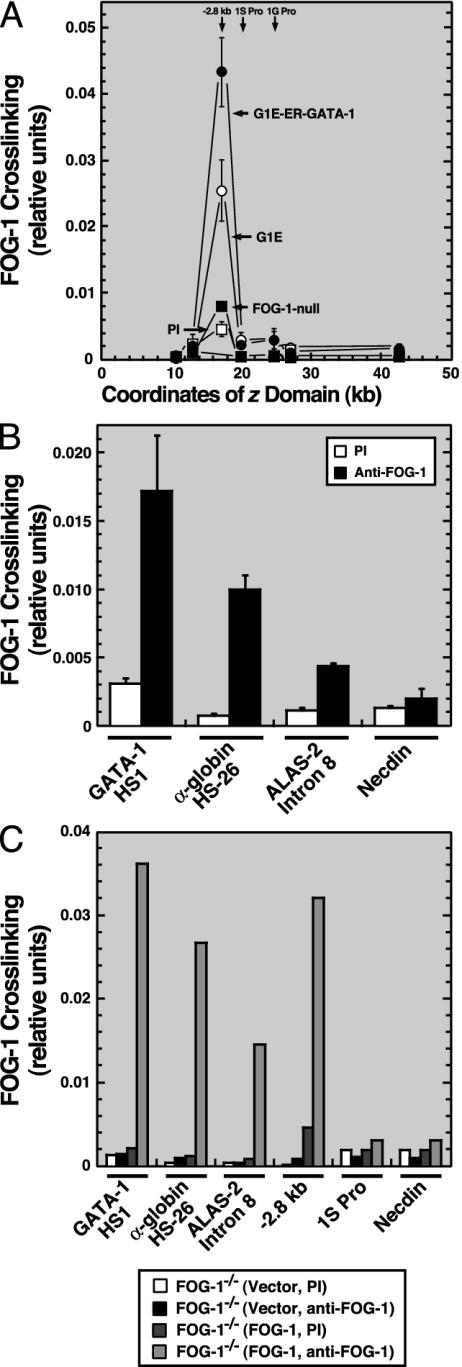
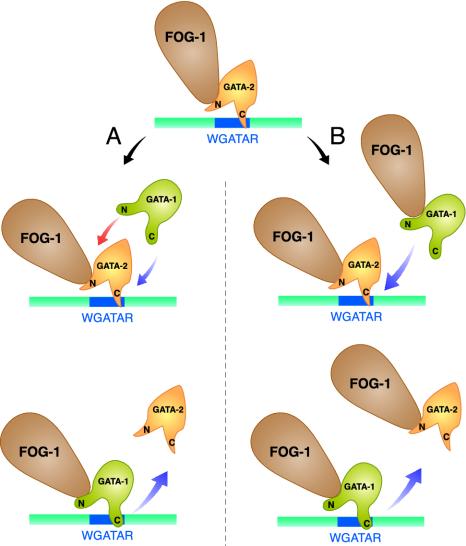
Coregulator recruitment by DNA-bound factors results in chromatin modification and protein-protein interactions, which regulate transcription. However, the mechanism by which the Friend of GATA (FOG) coregulator mediates GATA factor-dependent transcription is unknown. We showed previously that GATA-1 replaces GATA-2 at an upstream region of the GATA-2 locus, and that this GATA switch represses GATA-2. Genetic complementation analysis in FOG-1-null hematopoietic precursors revealed that FOG-1 is not required for establishment or maintenance of the active GATA-2 domain, but is critical for the GATA switch. Analysis of GATA factor binding to additional loci also revealed FOG-1-dependent GATA switches. Thus, FOG-1 facilitates chromatin occupancy by GATA-1 at sites bound by GATA-2. We propose that FOG-1 is a prototype of a new class of coregulators termed chromatin occupancy facilitators, which confer coregulation in certain contexts via enhancing trans-acting factor binding to chromatin in vivo.
Figures
References
-
- Brownell, J. E., Zhou, J., Ranalli, T., Kobayashi, R., Edmondson, D. G., Roth, S. Y. & Allis, C. D. (1996) Cell 84, 843–851. - PubMed
-
- Laherty, C. D., Yang, W. M., Sun, J. M., Davie, J. R., Seto, E. & Eisenman, R. N. (1997) Cell 89, 349–356. - PubMed
-
- Bresnick, E. H., Im, H. & Johnson, K. D. (2003) in Nature Encyclopedia of the Human Genome, ed. Cooper, D. N. (Nature Publishing Group, London), Vol. 3, pp. 260–264.
-
- Weiss, M. J. & Orkin, S. H. (1995) Exp. Hematol. 23, 99–107. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
