

Input-specific modulation of neurotransmitter release in the lateral horn of the spinal cord via adenosine receptors
- PMID: 14715946
- PMCID: PMC6729584
- DOI: 10.1523/JNEUROSCI.4591-03.2004
Input-specific modulation of neurotransmitter release in the lateral horn of the spinal cord via adenosine receptors
Abstract
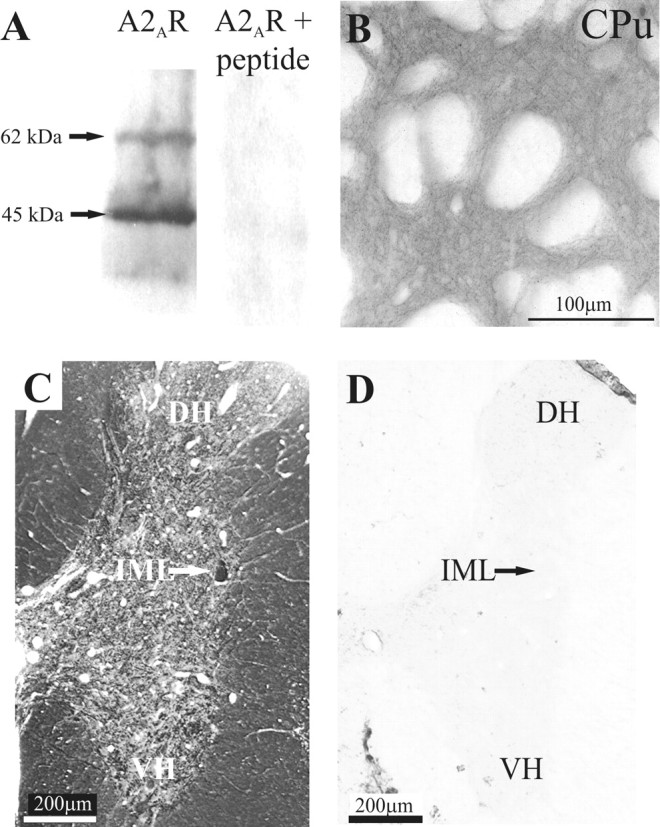
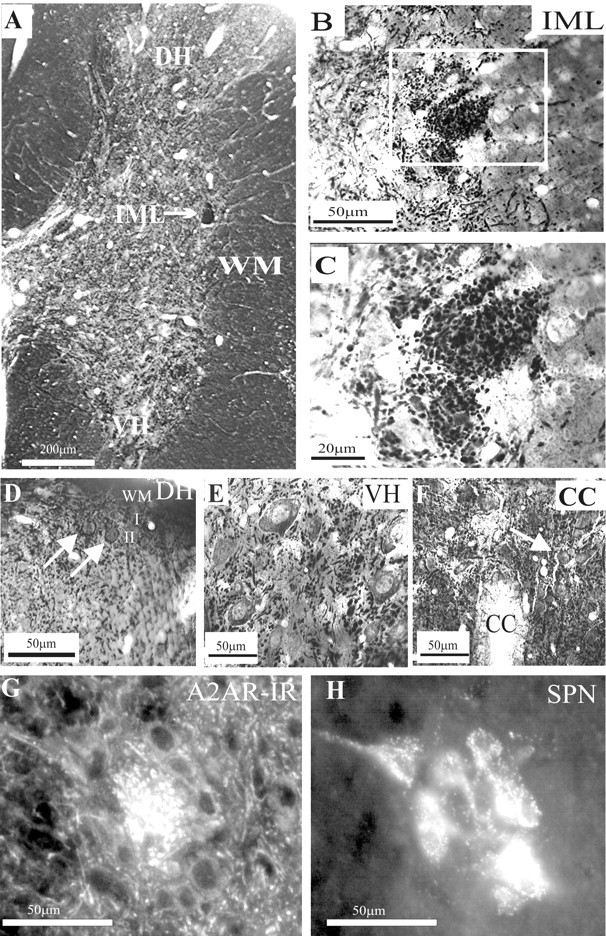
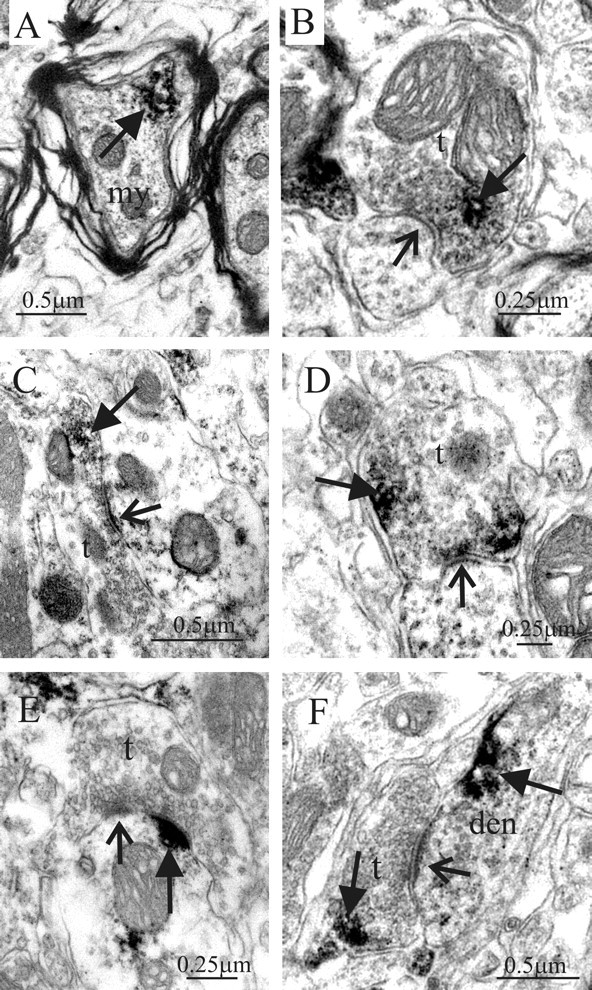
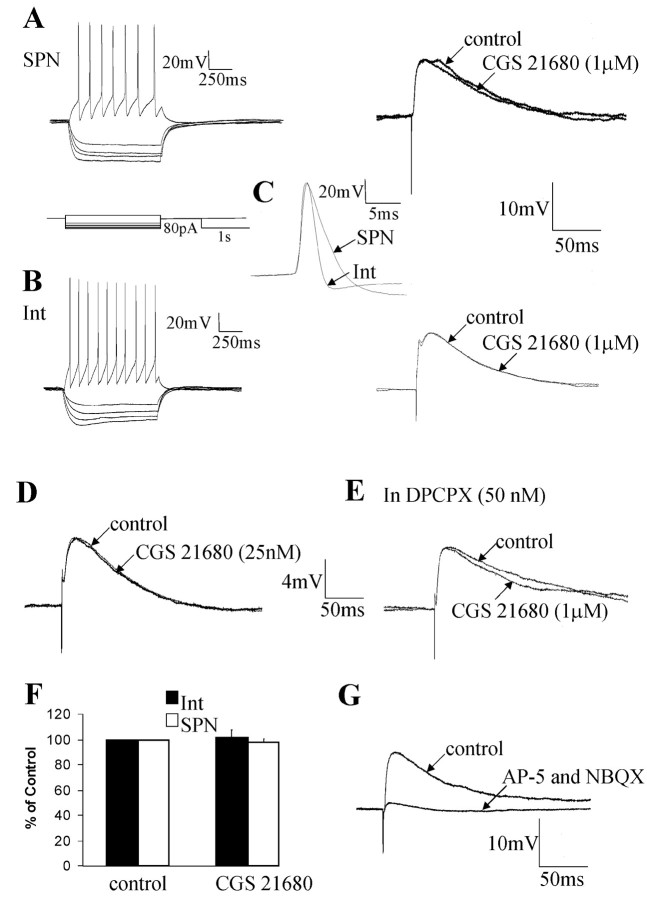
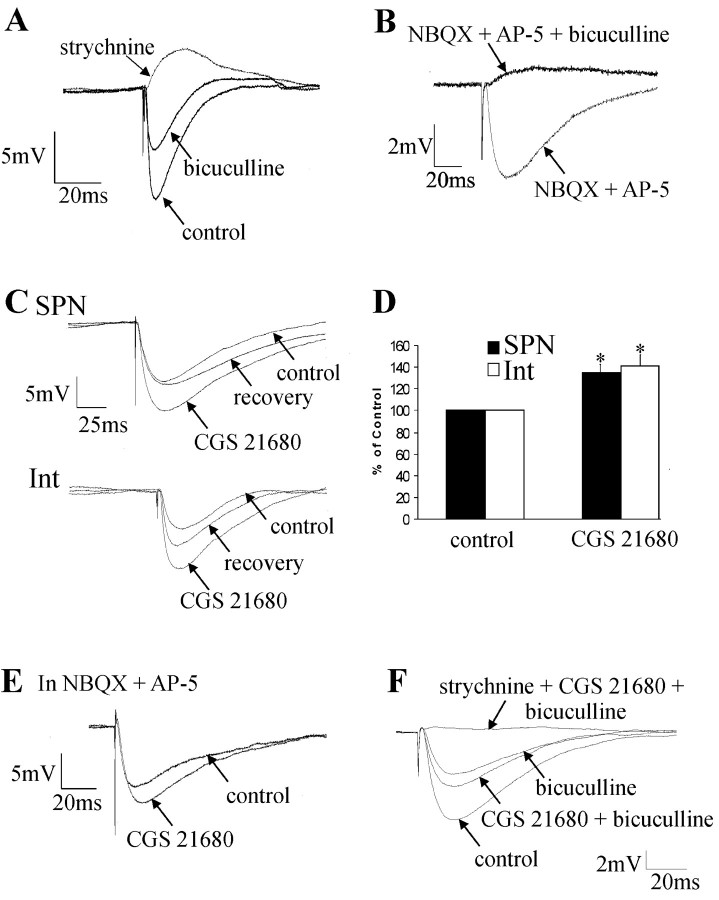
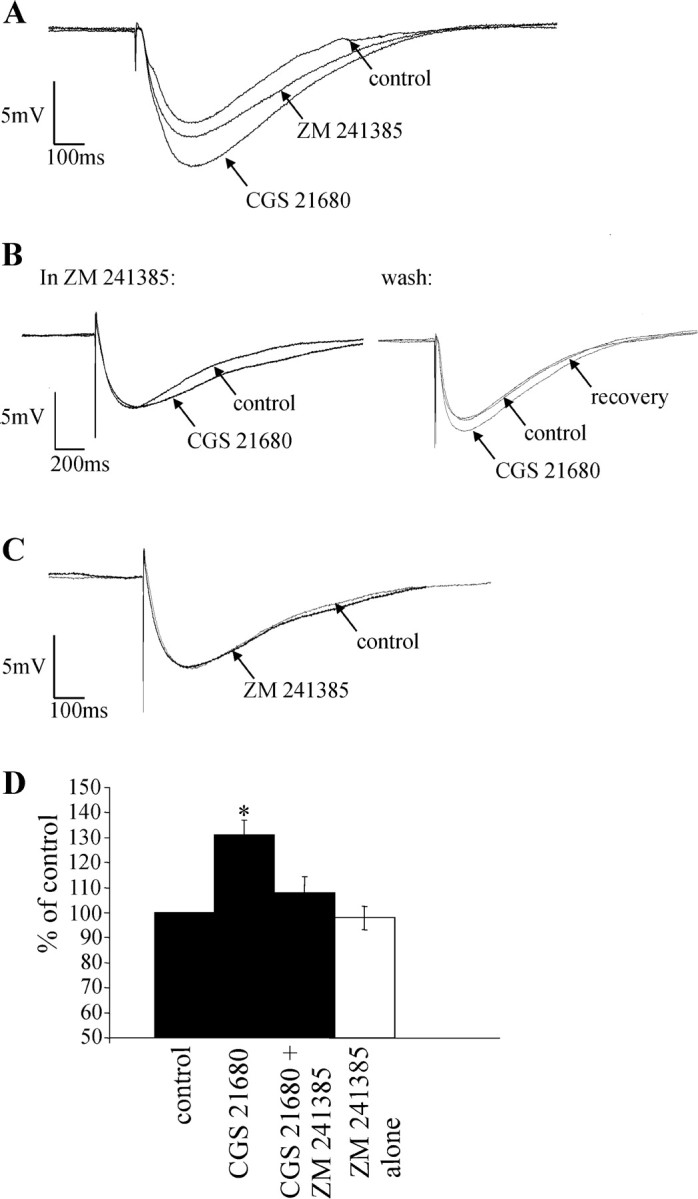
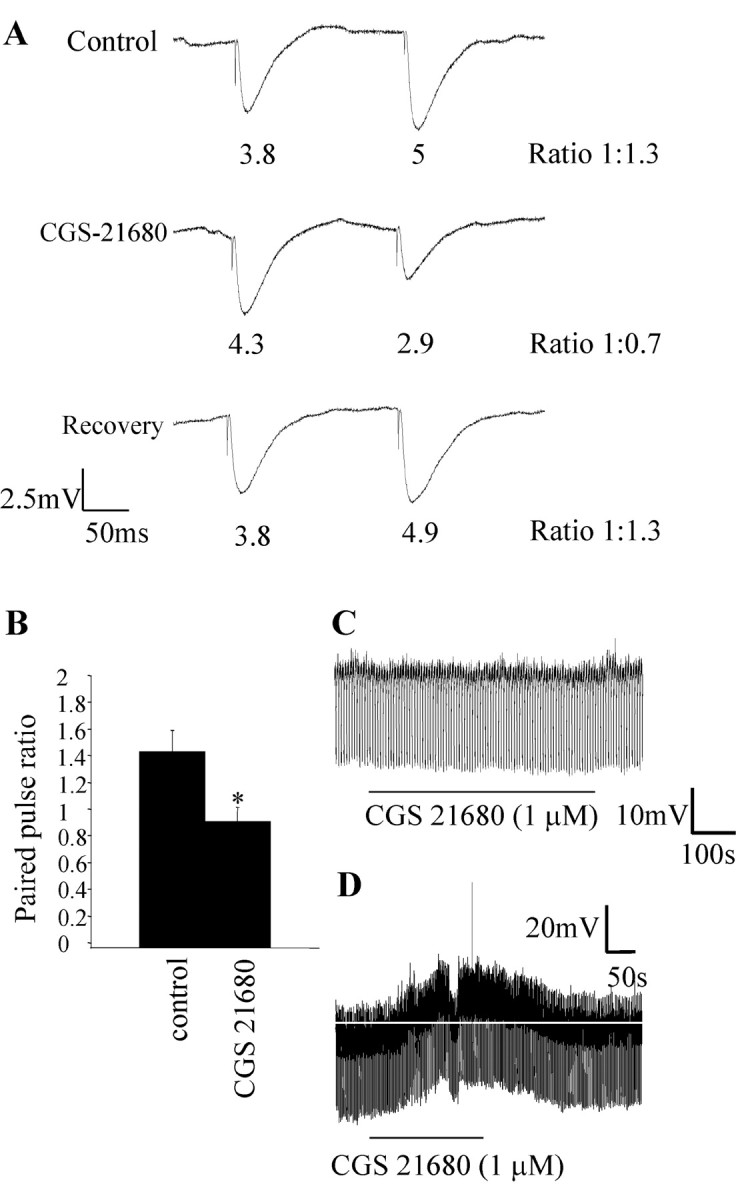
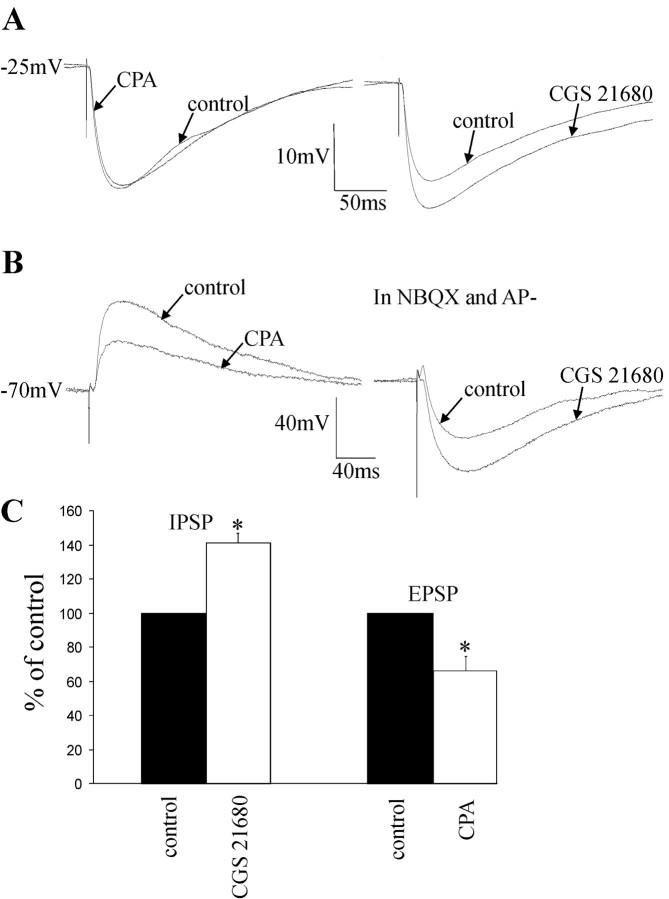
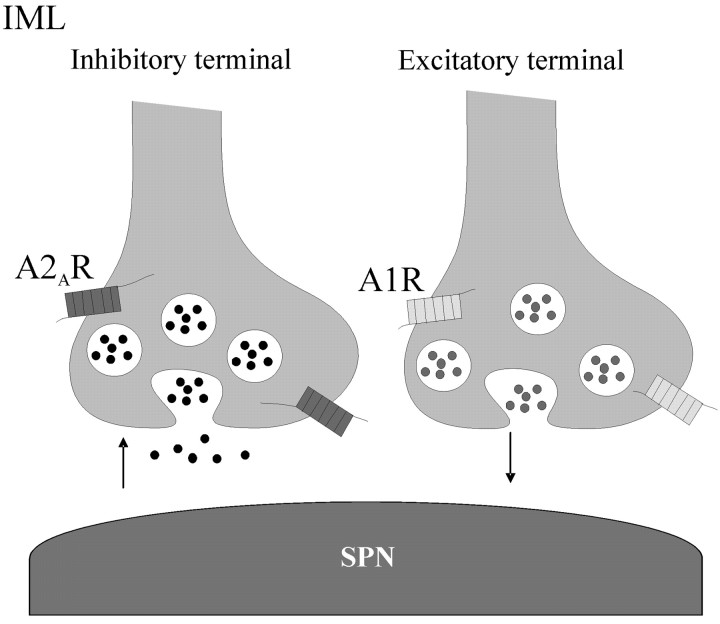
Activation of adenosine A2A receptors (A2ARs) in the CNS produces a variety of neuromodulatory actions dependent on the region and preparation examined. In autonomic regions of the spinal cord, A1R activation decreases excitatory synaptic transmission, but the effects of A2AR stimulation are unknown. We sought to determine the location and function of the A2ARs in the thoracic spinal cord, focusing on the intermediolateral cell column (IML). A2AR immunoreactivity was observed throughout the gray matter, with particularly dense immunostaining in regions containing sympathetic preganglionic neurons (SPNs), namely, the IML and intercalated nucleus. Electron microscopy revealed A2AR immunoreactivity within presynaptic terminals and in postsynaptic structures in the IML. To study the functional relevance of these A2ARs, visualized whole-cell patch-clamp recordings were made from electrophysiologically identified SPNs and interneurons within the IML. The A2AR agonist c2-[p-(carboxyethyl)phenethylamino]-5'-N-ethylcarboxyamidoadenosine (CGS 21680) had no significant effect on EPSPs but increased the amplitude of IPSPs elicited by stimulation of the lateral funiculus. These effects were attributable to activation of presynaptic A2ARs because CGS 21680 application altered the paired pulse ratio. Furthermore, neurons in the IML that have IPSPs increased via A2AR activation also receive excitatory inputs that are inhibited by A1R activation. These data show that activating A2ARs increase inhibitory but not excitatory transmission onto neurons in the IML. Simultaneous activation of A1Rs and A2ARs therefore could facilitate inhibition of the postsynaptic neuron, leading to an overall reduction of sympathetic nervous activity.
Figures
References
-
- Aden U, Halldner L, Lagercrantz H, Dalmau I, Ledent C, Fredholm BB (2003) Aggravated brain damage after hypoxic ischemia in immature adenosine A2A knockout mice. Stroke 34: 739-744. - PubMed
-
- Ambalavanar R, Morris R (1989) Fluoro-gold injected either subcutaneously or intravascularly results in extensive retrograde labelling of CNS neurones having axons terminating outside the blood-brain barrier. Brain Res 505: 171-175. - PubMed
-
- Anderson CR, Edwards SL (1994) Intraperitoneal injections of fluorogold reliably labels all sympathetic preganglionic neurons in the rat. J Neurosci Methods 53: 137-141. - PubMed
-
- Barraco RA, Martens KA, Parizon M, Normile HJ (1993) Adenosine A2a receptors in the nucleus accumbens mediate locomotor depression. Brain Res Bull 31: 397-404. - PubMed
-
- Blum D, Hourez R, Galas MC, Popoli P, Schiffmann SN (2003) Adenosine receptors and Huntington's disease: implications for pathogenesis and therapeutics. Lancet Neurol 2: 366-374. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous