

Ionic currents mediated by a prokaryotic homologue of CLC Cl- channels
- PMID: 14718478
- PMCID: PMC2217429
- DOI: 10.1085/jgp.200308935
Ionic currents mediated by a prokaryotic homologue of CLC Cl- channels
Abstract
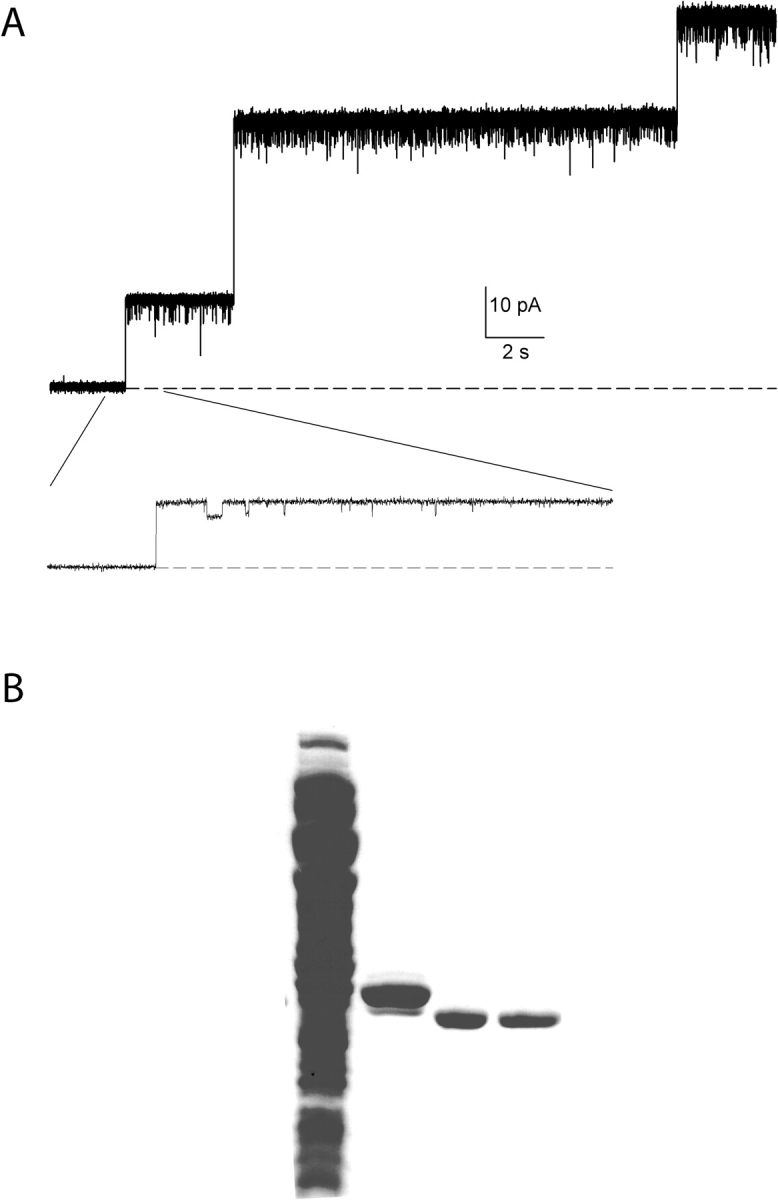
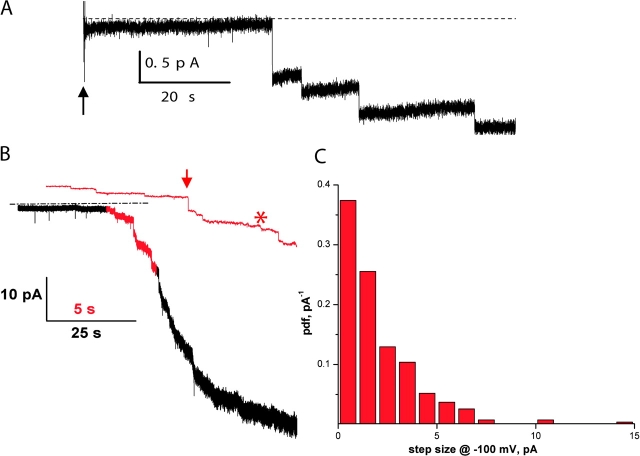
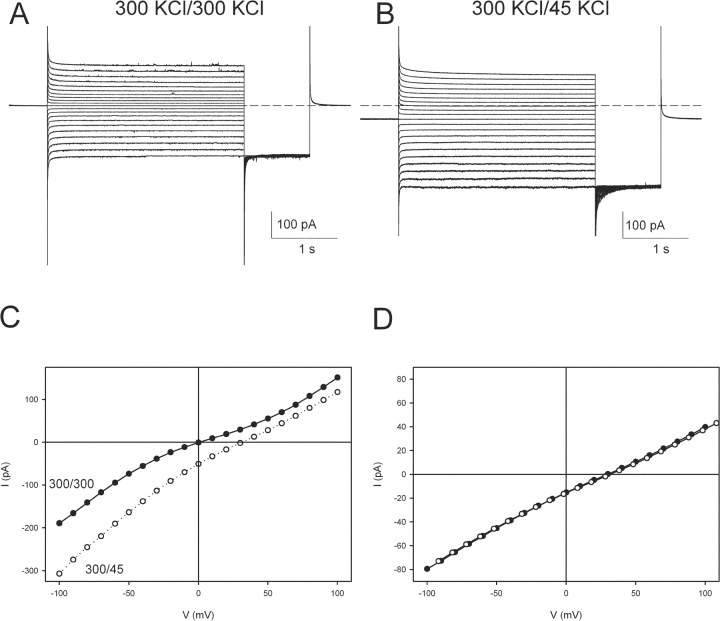
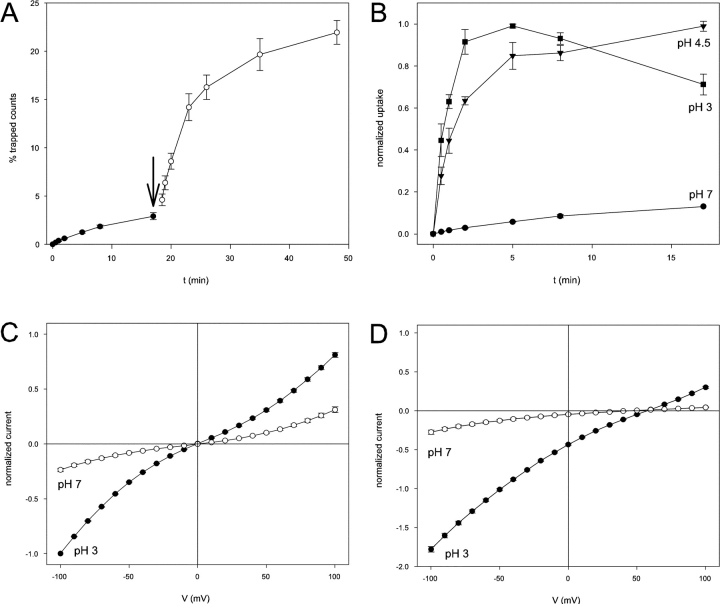
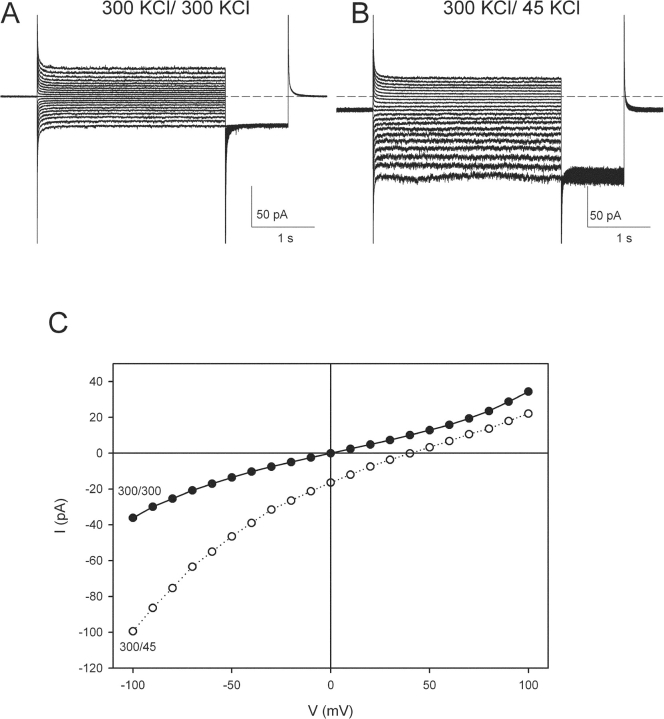
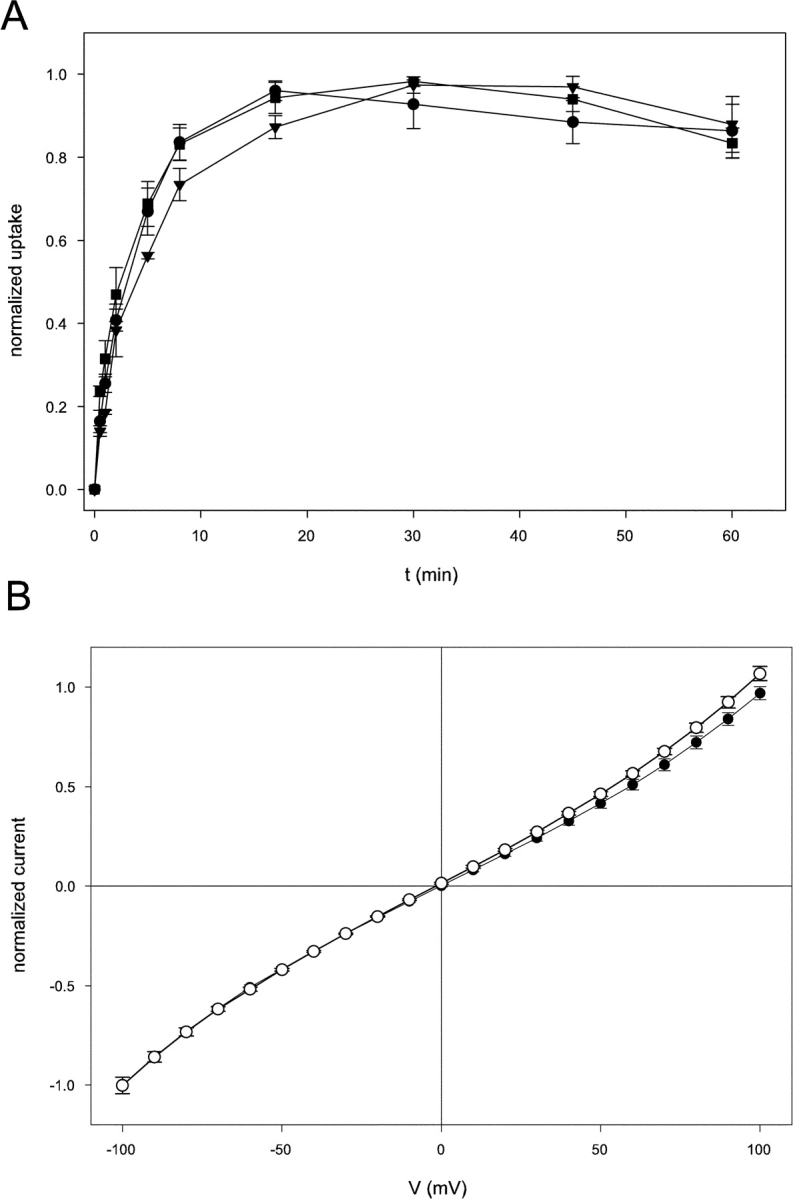
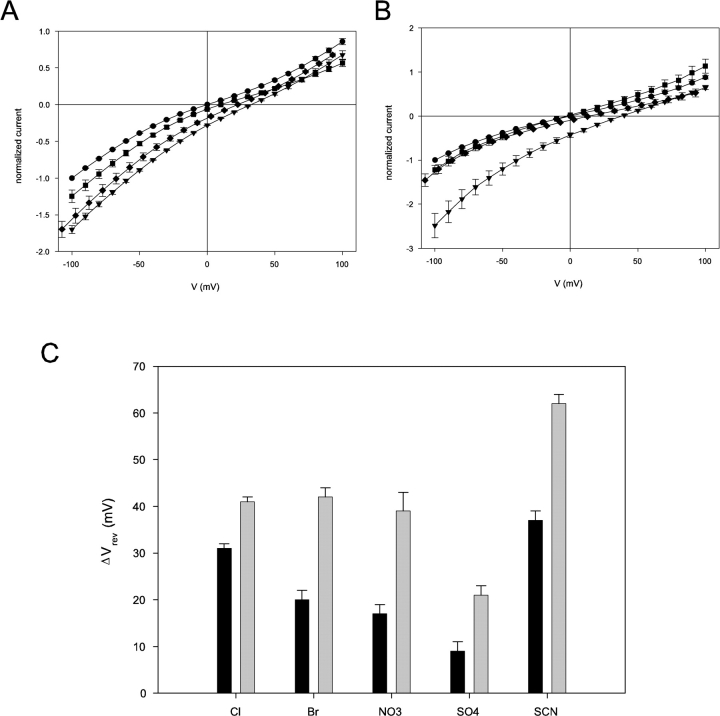
CLC-ec1 is an E. coli homologue of the CLC family of Cl- channels, which are widespread throughout eukaryotic organisms. The structure of this membrane protein is known, and its physiological role has been described, but our knowledge of its functional characteristics is severely limited by the absence of electrophysiological recordings. High-density reconstitution and incorporation of crystallization-quality CLC-ec1 in planar lipid bilayers failed to yield measurable CLC-ec1 currents due to porin contamination. A procedure developed to prepare the protein at a very high level of purity allowed us to measure macroscopic CLC-ec1 currents in lipid bilayers. The current is Cl- selective, and its pH dependence mimics that observed with a 36Cl- flux assay in reconstituted liposomes. The unitary conductance is estimated to be <0.2 pS. Surprisingly, the currents have a subnernstian reversal potential in a KCl gradient, indicating imperfect selectivity for anions over cations. Mutation of a conserved glutamate residue found in the selectivity filter eliminates the pH-dependence of both currents and 36Cl- flux and appears to trap CLC-ec1 in a constitutively active state. These effects correlate well with known characteristics of eukaryotic CLC channels. The E148A mutant displays nearly ideal Cl- selectivity.
Figures
References
-
- Benz, R., and K. Bauer. 1988. Permeation of hydrophilic molecules through the outer membrane of gram-negative bacteria. Review on bacterial porins. Eur. J. Biochem. 176:1–19. - PubMed
-
- Dutzler, R., E.B. Campbell, M. Cadene, B.T. Chait, and R. MacKinnon. 2002. X-ray structure of a ClC chloride channel at 3.0 Å reveals the molecular basis of anion selectivity. Nature. 415:287–294. - PubMed
-
- Dutzler, R., E.B. Campbell, and R. MacKinnon. 2003. Gating the selectivity filter in ClC chloride channels. Science. 300:108–112. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
