

doi: 10.1084/jem.20031615.
Epub 2004 Jan 12.
Beta-catenin is dispensable for hematopoiesis and lymphopoiesis
Affiliations
- PMID: 14718516
- PMCID: PMC2211763
- DOI: 10.1084/jem.20031615
Item in Clipboard
Beta-catenin is dispensable for hematopoiesis and lymphopoiesis
J Exp Med.
.
Abstract
Beta-catenin-mediated Wnt signaling has been suggested to be critically involved in hematopoietic stem cell maintenance and development of T and B cells in the immune system. Unexpectedly, here we report that inducible Cre-loxP-mediated inactivation of the beta-catenin gene in bone marrow progenitors does not impair their ability to self-renew and reconstitute all hematopoietic lineages (myeloid, erythroid, and lymphoid), even in competitive mixed chimeras. In addition, both thymocyte survival and antigen-induced proliferation of peripheral T cells is beta-catenin independent. In contrast to earlier reports, these data exclude an essential role for beta-catenin during hematopoiesis and lymphopoiesis.
Figures
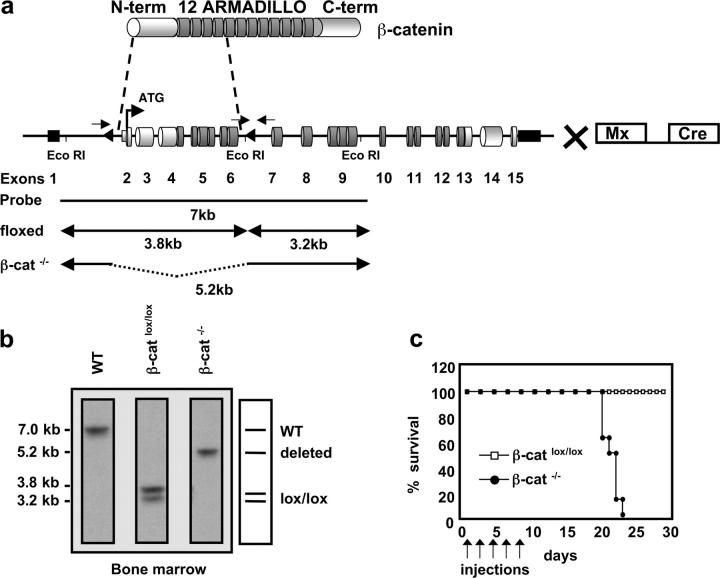
Inducible targeting of the β-catenin gene in the BM by the Mx-Cre recombinase. (a) Schematic representation of the β-catenin protein containing an NH2-terminal region (N-term), 12 Armadillo repeats, and a carboxy terminus harboring the transactivation domain (C-term). The genomic organization of the β-cateninlox/lox locus is shown with cylinders indicating coding exons and small black rectangles indicating noncoding exons. Exons 2–6 are flanked by two loxP sequences (triangles). β-cateninlox/lox mice were crossed to mice carrying the IFN-α–inducible Mx-Cre transgene to inducibly inactivate β-catenin in the BM after pI-pC treatment. Arrows indicate approximate positions of the PCR primers used to verify the deletion of the floxed β-catenin locus. EcoRI restriction sites and the probe for Southern blot analysis are indicated. (b) Southern blot analysis of Eco-RI–digested genomic DNA from BM of WT, β-cateninlox/lox, or β-catenin−/− mice. The probe indicated in (a) reveals a 7-kb fragment for the WT allele, two fragments of 3.8 and 3.2 kb for the homozygously floxed β-catenin locus, and one 5.2-kb fragment for the β-catenin−/− allele after successful deletion of the loxP flanked gene segment. (c) Survival curve of β-cateninlox/lox and β-catenin−/− mice after injection of pI-pC as indicated by arrows. Death of β-catenin−/− mice starts 2 wk after the last injection. n = 8 for both β-cateninlox/lox (controls) and β-catenin−/−.
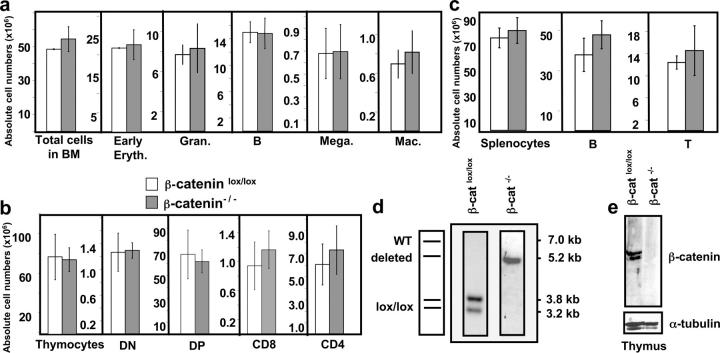
β-catenin–deficient BM reconstitutes all major hematopoietic lineages. BM chimeras were analyzed 4–7 mo after reconstitution. (a) Absolute donor cell numbers for total BM and different subsets: early erythroblasts (Early Eryth., Ter119+), granulocytes (Gran., Gr1+), B cells (B220+), megakaryocytes (Mega., CD41+), and macrophages (Mac, Gr1− CD11b+) of either β-cateninlox/lox (open bars) or β-catenin−/− (shaded bars) mice. The bars represent mean ± SD values (n = 5). (b) Absolute donor cell numbers for total thymocytes and thymocyte subsets: CD4− CD8− (DN), CD4+ CD8+ (DP), CD4+ CD8− TCRβ+ (CD4), and CD8+ CD4− TCRβ+ (CD8). The bars represent mean ± SD values for β-cateninlox/lox (open bars) or β-catenin−/− (filled bars) mice, where n = 5 for both β-cateninlox/lox and β-catenin−/− mice. (c) Absolute donor cell numbers for total splenocytes, B cells (B220+), and T cells (CD3+). n = 5 for control and β-catenin−/− mice. (d) Southern blot analysis of Eco-RI–digested genomic DNA from donor thymocytes of control (β-cateninlox/lox) or β-catenin−/− BM chimeric mice (refer to Fig.1 a for details). (e) Immunoblot analysis of β-catenin protein expression in total donor thymocytes from control or β-catenin−/− BM chimeras. The blots were probed with an antiserum specific to the COOH terminus of the β-catenin protein. To verify that equal amounts of protein were loaded, the membrane was reprobed with a monoclonal antibody against α-tubulin (bottom).
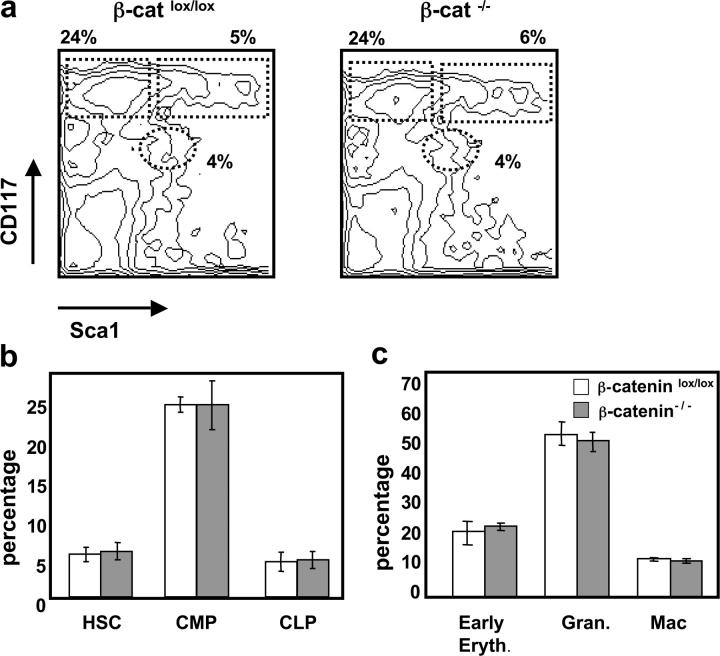
Normal development of β-catenin−/− hematopoietic lineages in a competitive situation. Mixed BM chimeric mice were analyzed 4–6 mo after reconstitution with a 1:2 mixture of WT (CD45.1+) and either β-cateninlox/lox or β-catenin−/− (CD45.2+) BM-derived populations. (a) A representative FACS® analysis of BM stained with anti-CD117 and anti-Sca1 gated on lin− CD45.2+ donor cells of either β-cateninlox/lox or β-catenin−/− mixed chimeras. Numbers indicate relative percentage of HSCs (CD117+ Sca1+) CLPs (CD117low Sca1low), and CMPs (CD117+ Sca1−) in each indicated region. (b) Percentage HSCs, CMPs, and CLPs gated on lin− CD45.2+ cells from either β-cateninlox/lox (open bars) or β-catenin−/− mixed chimeras (shaded bars). The bars represent mean ± SD (n = 8). (c) Percentage early erythroblasts (Ter119+), granulocytes (Gr1+), and macrophages (Gr1− CD11b+) after gating on CD45.2+ cells from either β-cateninlox/lox or β-catenin−/− mixed chimeras. The bars represent mean ± SD (n = 8).
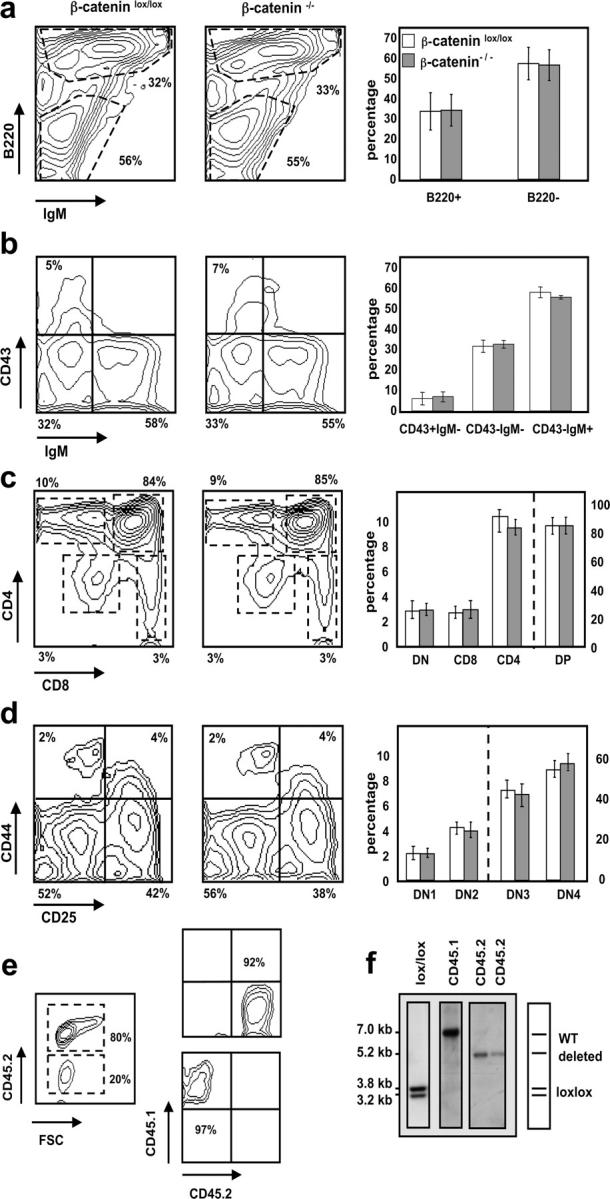
Normal development of β-catenin–deficient B and T cells in mixed BM chimeras. Representative FACS® analyses (left and middle panels) gated on CD45.2+ donor cells. Bar diagrams (right panels) indicate relative number ± SD of the different subsets where n = 12 for both β-cateninlox/lox (open bars) and β-catenin−/− (shaded bars) mice. (a) BM stained with anti-B220 and anti-IgM antibodies. (b) BM cells stained with anti-CD43 and anti-IgM antibodies after gating on B220+ cells. (c) Total thymocytes stained with anti-CD4 and anti-CD8 antibodies. DN (CD4− CD8−), DP (CD4+ CD8+), CD4 (CD4+ CD8− TCRβ+), and CD8 (CD4− CD8+ TCRβ+). (d) Thymocytes stained with anti-CD44 and anti-CD25 antibodies after gating on donor (CD45.2+)-derived lineage-negative cells. DN1 (CD44+ CD25−), DN2 (CD44+ CD25+), DN3 (CD44− CD25+), and DN4 (CD44− CD25−). (e) CD45.2+ (donor) and CD45.1+ (WT) cells derived from chimeric spleens were purified by FACS® sorting to verify deletion of the floxed β-catenin alleles after reconstitution. FACS® analysis before (left) and after the sort (right). Relative purity of the sorted populations are indicated. (f) Southern blot analysis of Eco-RI–digested genomic DNA from sorted CD45.1+ and CD45.2+ donor splenocytes. CD45.1+ cells were pooled from three different mice and CD45.2+ cells are derived from two individual mice.
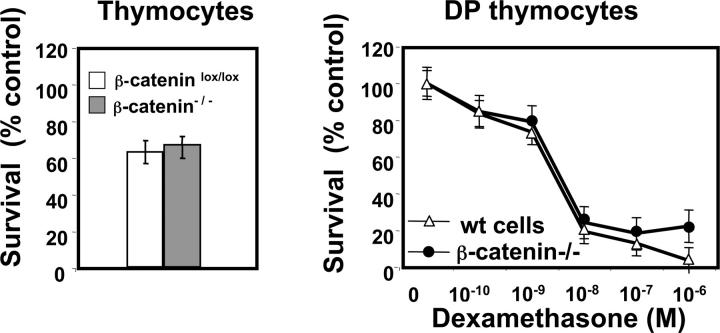
Absence of β-catenin does not affect spontaneous or glucocorticoid-induced death of thymocytes. (a) Percentage of surviving thymocytes gated on CD45.1+ (WT donor) or CD45.2+ (β-catenin−/− donor) derived from mixed BM chimeras after 12 h of culture at 37°C. The percentage of surviving cells is normalized to control cells maintained at 4°C. Data represent mean ± SD of five experiments. (b) Glucocorticoid-induced cell death in WT (CD45.1+) and β-catenin−/− (CD45.2+) CD4+ CD8+ DP thymocytes after 12 h of culture at 37°C. Data are normalized to untreated controls and represent mean ± SD from five individual experiments.
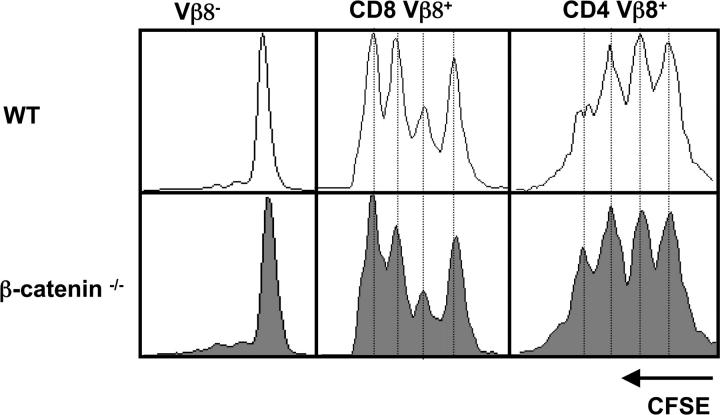
Loss of β-catenin does not perturb antigen-induced T cell proliferation. CFSE-labeled splenocytes isolated from mixed BM chimeras were transferred i.v. into WT mice. The next day mice were injected i.p. with 20 μg SEB. 2 d later the draining popliteal LNs were removed and analyzed by FACS®. After gating on CD45.1+ (WT donor) or CD45.2+ (β-catenin−/− donor), the number of cell divisions in Vβ8− (negative control), CD8+ Vβ8+, and CD4+ Vβ8+ populations are shown. One representative experiment out of four is shown.
References
-
- Morrison, S.J., N. Uchida, and I.L. Weissman. 1995. The biology of hematopoietic stem cells. Annu. Rev. Cell Dev. Biol. 11:35–71. - PubMed
-
- van de Wetering, M., W. de Lau, and H. Clevers. 2002. WNT signaling and lymphocyte development. Cell. 109:S13–S19. - PubMed
-
- Reya, T., A.W. Duncan, L. Ailles, J. Domen, D.C. Scherer, K. Willert, L. Hintz, R. Nusse, and I.L. Weissman. 2003. A role for Wnt signalling in self-renewal of haematopoietic stem cells. Nature. 423:409–414. - PubMed
-
- Willert, K., J.D. Brown, E. Danenberg, A.W. Duncan, I.L. Weissman, T. Reya, J.R. Yates III, and R. Nusse. 2003. Wnt proteins are lipid-modified and can act as stem cell growth factors. Nature. 423:448–452. - PubMed
-
- Bhanot, P., M. Brink, C.H. Samos, J.C. Hsieh, Y. Wang, J.P. Macke, D. Andrew, J. Nathans, and R. Nusse. 1996. A new member of the frizzled family from Drosophila functions as a Wingless receptor. Nature. 382:225–230. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
