

ARC-1, a sequence element complementary to an internal 18S rRNA segment, enhances translation efficiency in plants when present in the leader or intercistronic region of mRNAs
- PMID: 14718549
- PMCID: PMC373286
- DOI: 10.1093/nar/gkh176
ARC-1, a sequence element complementary to an internal 18S rRNA segment, enhances translation efficiency in plants when present in the leader or intercistronic region of mRNAs
Abstract
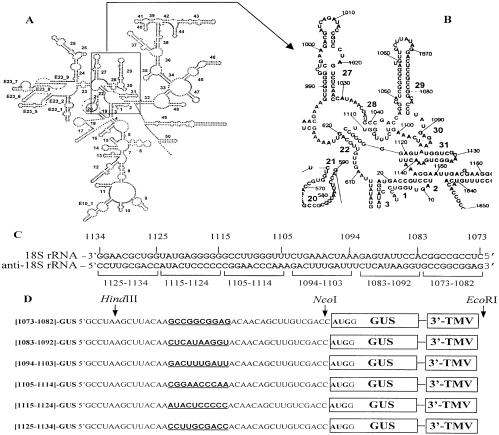
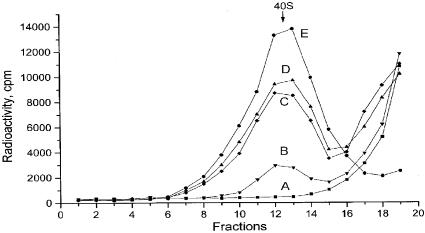
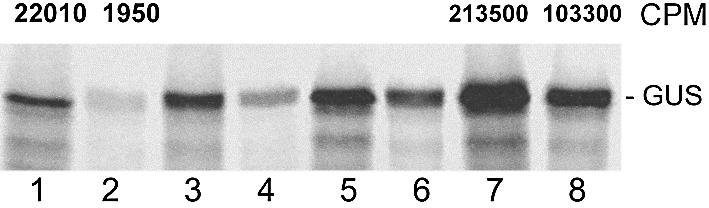
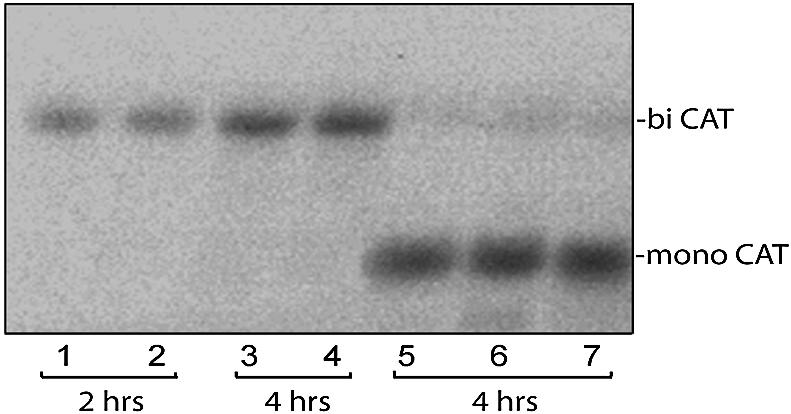
The sequences of different plant viral leaders with known translation enhancer ability show partial complementarity to the central region of 18S rRNA. Such complementarity might serve as a means to attract 40S ribosomal subunits and explain in part the translation-enhancing property of these sequences. To verify this notion, we designed beta-glucuronidase (GUS) mRNAs differing only in the nature of 10 nt inserts in the center of their 41 base leaders. These were complementary to consecutive domains of plant 18S rRNA. Sucrose gradient analysis revealed that leaders with inserts complementary to regions 1105-1114 and 1115-1124 ('ARC-1') of plant 18S rRNA bound most efficiently to the 40S ribosomal subunit after dissociation from 80S ribosomes under conditions of high ionic strength, a treatment known to remove translation initiation factors. Using wheat germ cell-free extracts, we could demonstrate that mRNAs with these leaders were translated more than three times more efficiently than a control lacking such a complementarity. Three linked copies of the insert enhanced translation of reporter mRNA to levels comparable with those directed by the natural translation enhancing leaders of tobacco mosaic virus and potato virus Y RNAs. Moreover, inserting the same leaders as intercistronic sequences in dicistronic mRNAs substantially increased translation of the second cistron, thereby revealing internal ribosome entry site activity. Thus, for plant systems, the complementary interaction between mRNA leader and the central region of 18S rRNA allows cap-independent binding of mRNA to the 43S pre-initiation complex without assistance of translation initiation factors.
Figures
References
-
- Gingras A.-C., Raught,B. and Sonenberg,N. (1999) eIF4 initiation factors: effectors of mRNA recruitment to ribosomes and regulators of translation. Annu. Rev. Biochem., 68, 913–963. - PubMed
-
- Gallie D.R. (1996) Translational control of cellular and viral mRNAs. Plant Mol. Biol., 32, 145–158. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
