

Two modes of polyamine block regulating the cardiac inward rectifier K+ current IK1 as revealed by a study of the Kir2.1 channel expressed in a human cell line
- PMID: 14724206
- PMCID: PMC1664885
- DOI: 10.1113/jphysiol.2003.055434
Two modes of polyamine block regulating the cardiac inward rectifier K+ current IK1 as revealed by a study of the Kir2.1 channel expressed in a human cell line
Abstract
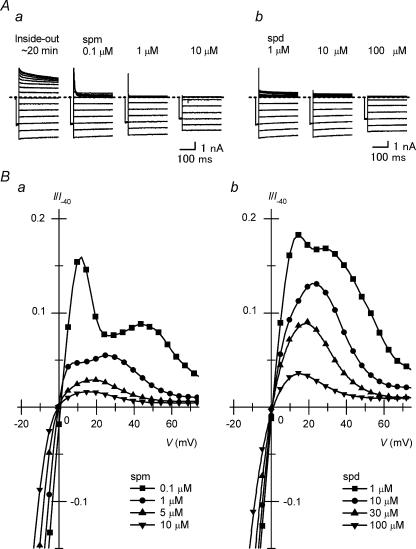
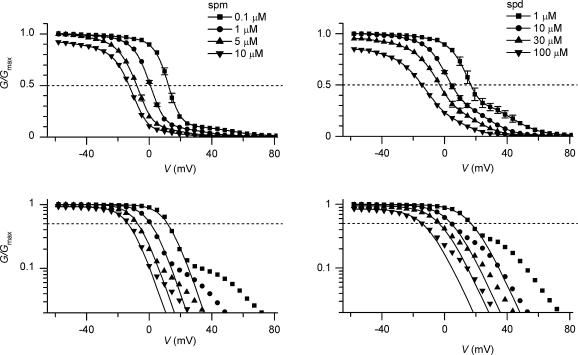
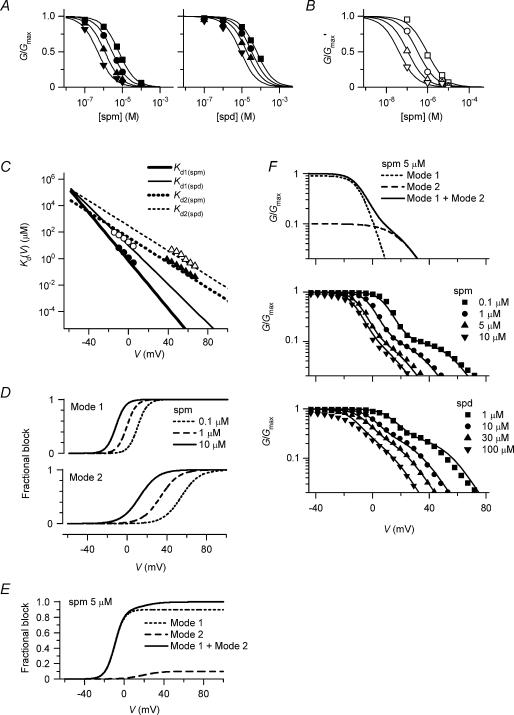
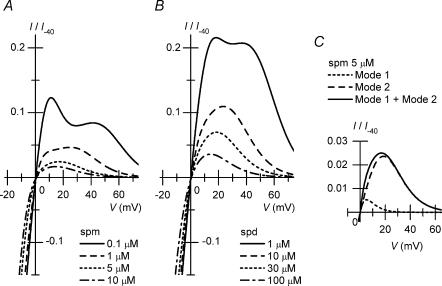
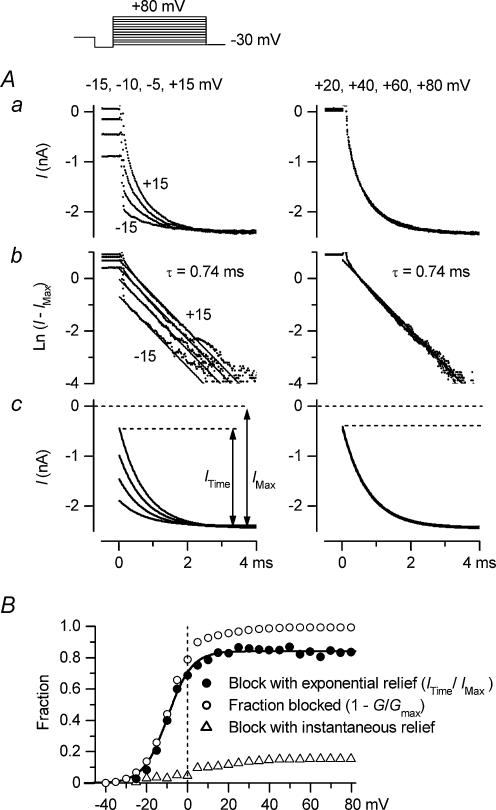
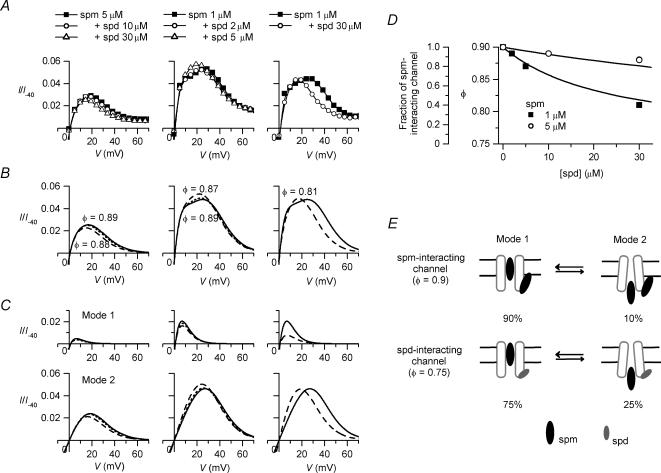
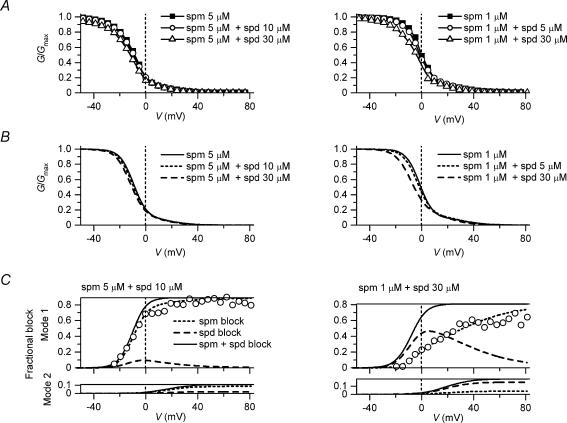
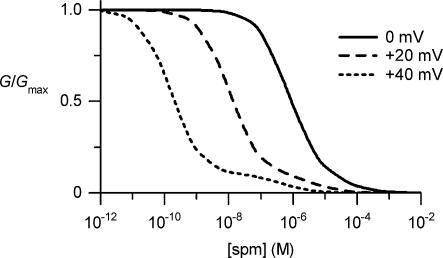
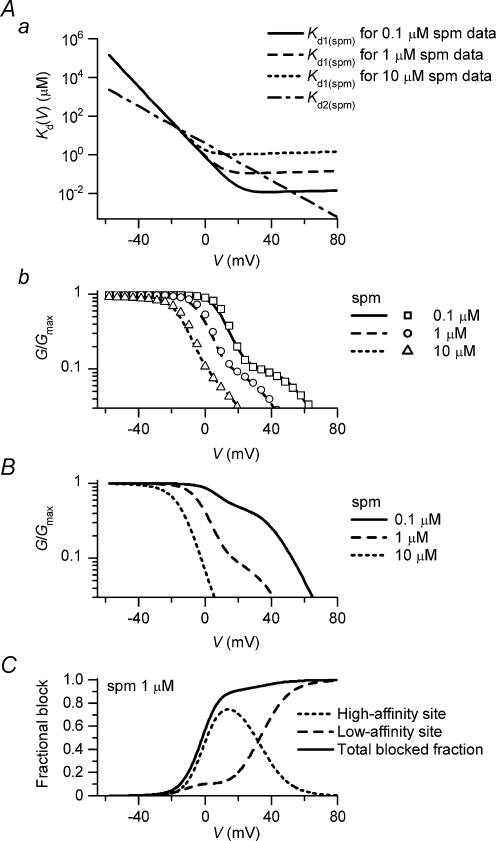
The strong inward rectifier K(+) current, I(K1), shows significant outward current amplitude in the voltage range near the reversal potential and thereby causes rapid repolarization at the final phase of cardiac action potentials. However, the mechanism that generates the outward I(K1) is not well understood. We recorded currents from the inside-out patches of HEK 293T cells that express the strong inward rectifier K(+) channel Kir2.1 and studied the blockage of the currents caused by cytoplasmic polyamines, namely, spermine and spermidine. The outward current-voltage (I-V) relationships of Kir2.1, obtained with 5-10 microm spermine or 10-100 microm spermidine, were similar to the steady-state outward I-V relationship of I(K1), showing a peak at a level that is approximately 20 mV more positive than the reversal potential, with a negative slope at more positive voltages. The relationships exhibited a plateau or a double-hump shape with 1 microm spermine/spermidine or 0.1 microm spermine, respectively. In the chord conductance-voltage relationships, there were extra conductances in the positive voltage range, which could not be described by the Boltzmann relations fitting the major part of the relationships. The extra conductances, which generated most of the outward currents in the presence of 5-10 microm spermine or 10-100 microm spermidine, were quantitatively explained by a model that considered two populations of Kir2.1 channels, which were blocked by polyamines in either a high-affinity mode (Mode 1 channel) or a low-affinity mode (Mode 2 channel). Analysis of the inward tail currents following test pulses indicated that the relief from the spermine block of Kir2.1 consisted of an exponential component and a virtually instantaneous component. The fractions of the two components nearly agreed with the fractions of the blockages in Mode 1 and Mode 2 calculated by the model. The estimated proportion of Mode 1 channels to total channels was 0.9 with 0.1-10 microm spermine, 0.75 with 1-100 microm spermidine, and between 0.75 and 0.9 when spermine and spermidine coexisted. An interaction of spermine/spermidine with the channel at an intracellular site appeared to modify the equilibrium of the two conformational channel states that allow different modes of blockage. Our results suggest that the outward I(K1) is primarily generated by channels with lower affinities for polyamines. Polyamines may regulate the amplitude of the outward I(K1), not only by blocking the channels but also by modifying the proportion of channels that show different sensitivities to the polyamine block.
Figures
References
-
- Fabiato A, Fabiato F. Calculator programs for computing the composition of the solutions containing multiple metals and ligands used for experiments in skinned muscle cells. J Physiol Paris. 1979;75:463–505. - PubMed
-
- Fakler B, Brandle U, Glowatzki E, Weidemann S, Zenner H-P, Ruppersberg JP. Strong voltage-dependent inward rectification of inward rectifier K+ channels is caused by intracellular spermine. Cell. 1995;80:149–154. - PubMed
-
- Ficker E, Taglialatela M, Wible BA, Henley CM, Brown AM. Spermine and spermidine as gating molecules for inward rectifier K+ channels. Science. 1994;266:1068–1072. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous