

Keratan sulfate proteoglycan phosphacan regulates mossy fiber outgrowth and regeneration
- PMID: 14724244
- PMCID: PMC6729989
- DOI: 10.1523/JNEUROSCI.3040-03.2004
Keratan sulfate proteoglycan phosphacan regulates mossy fiber outgrowth and regeneration
Abstract
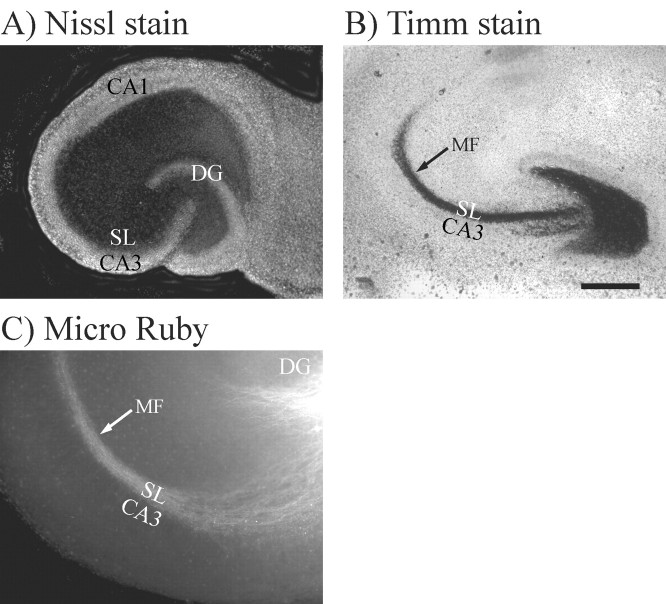
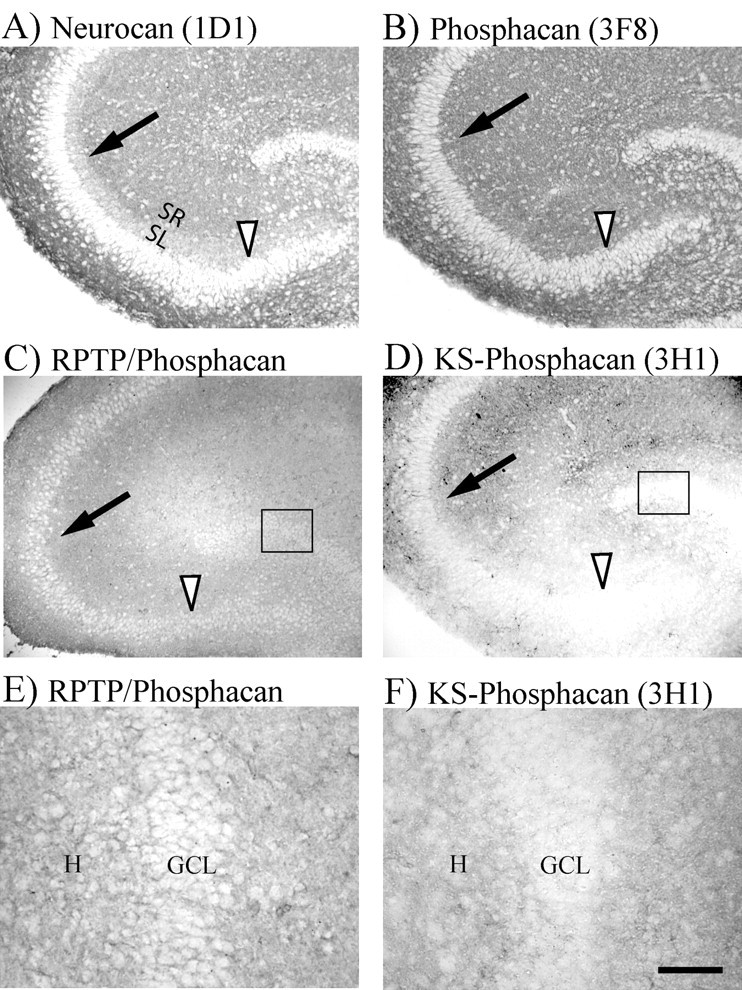
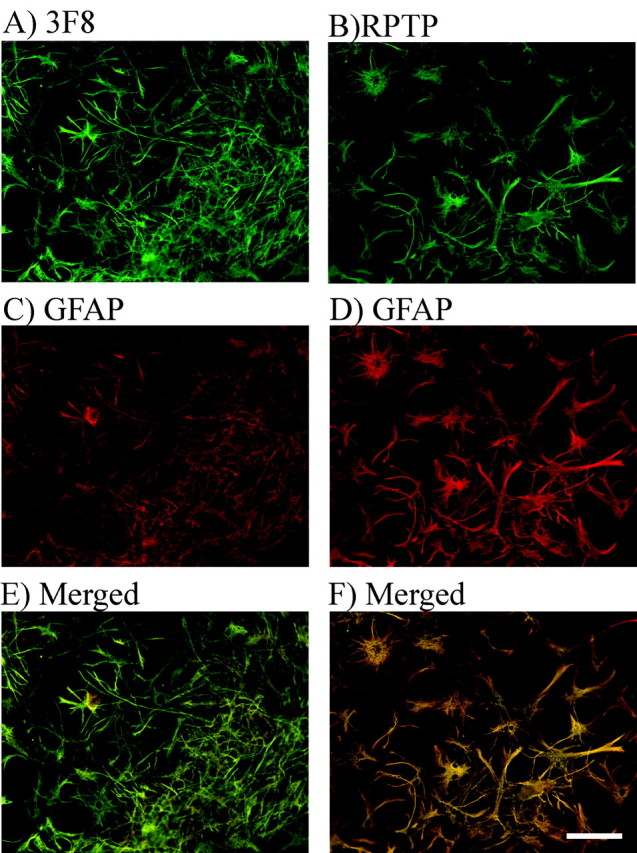
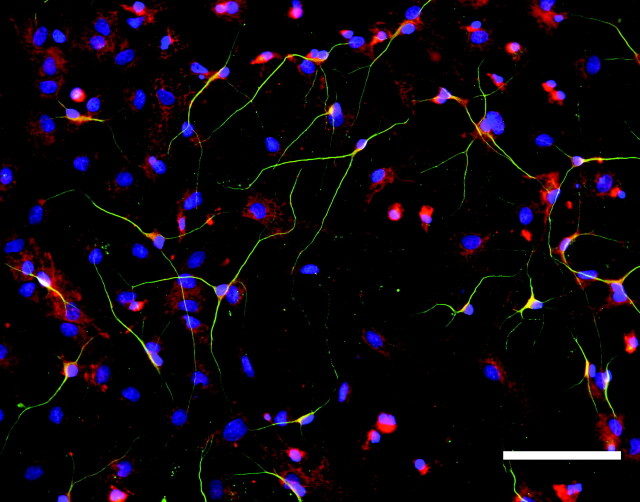
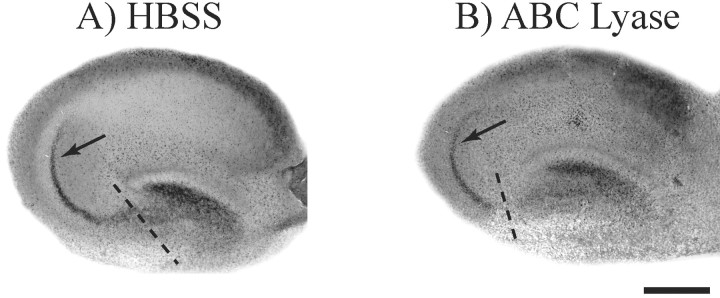
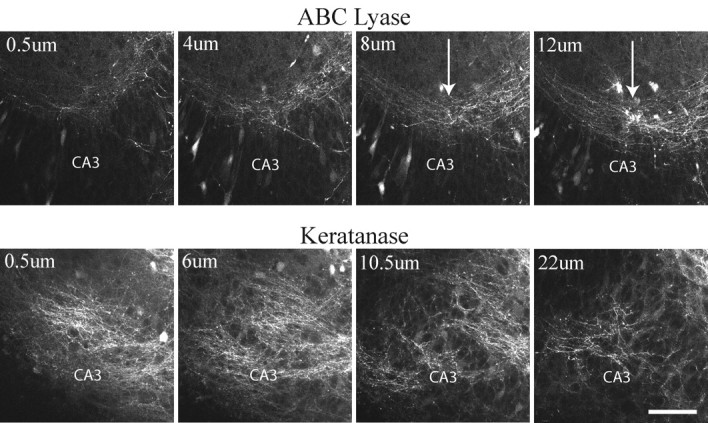
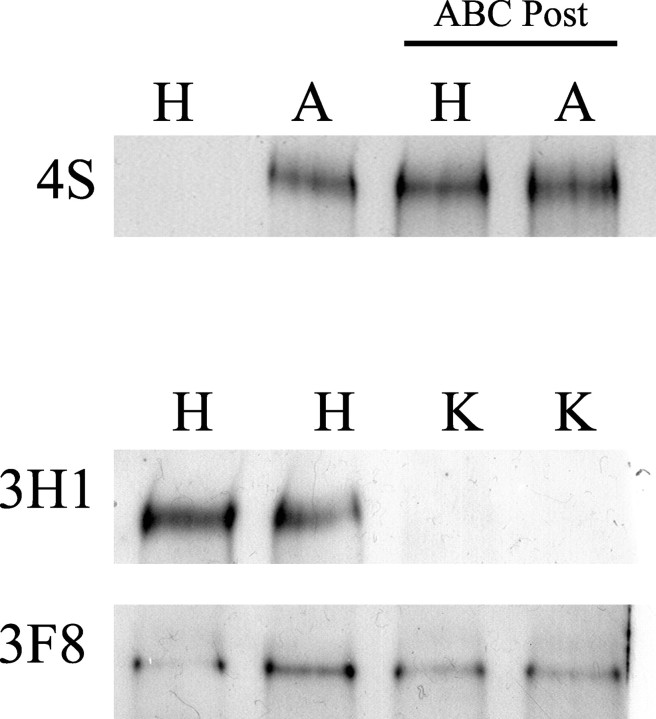
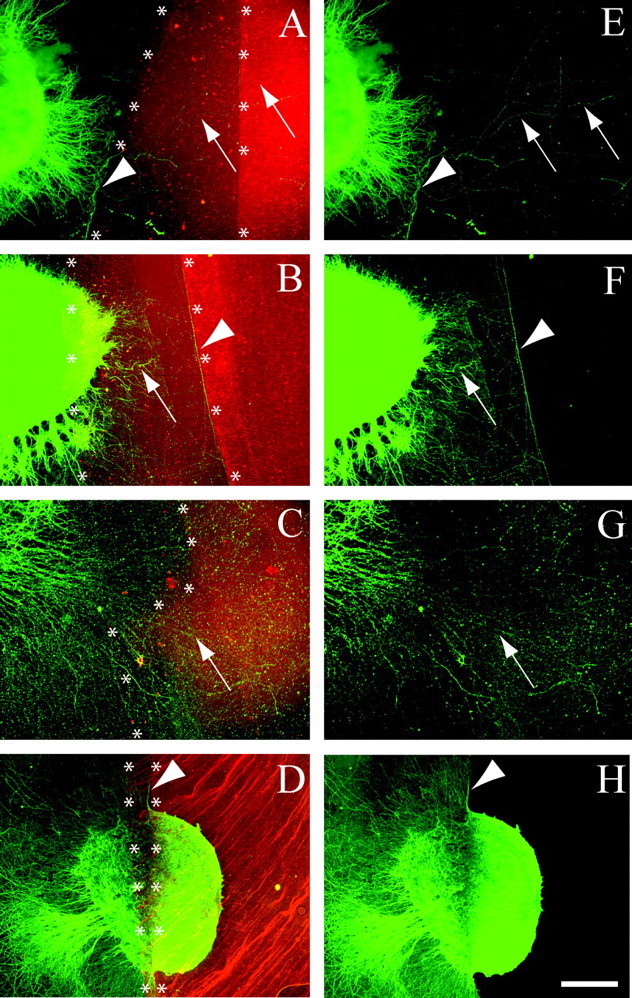
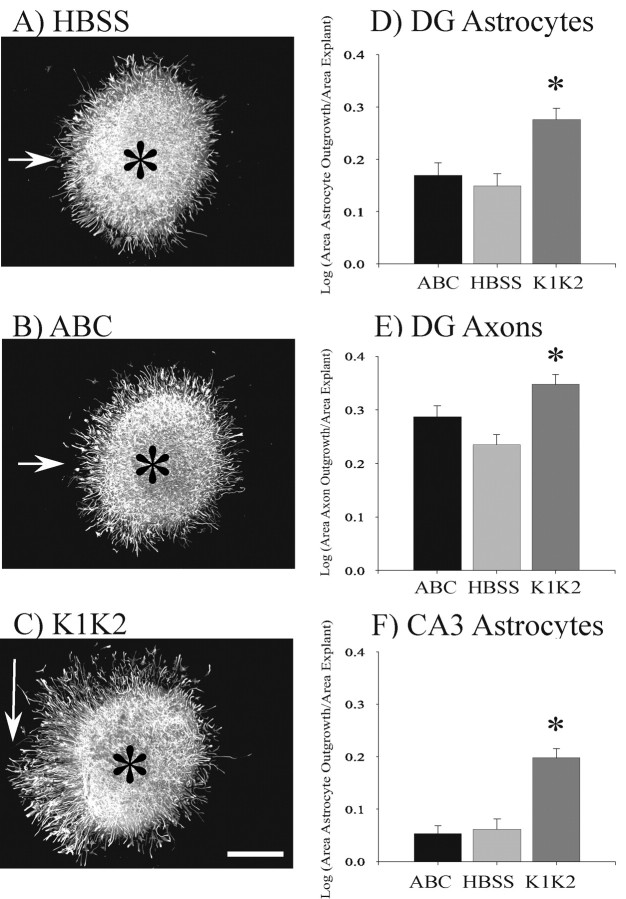
We have examined the role of chondroitin sulfate proteoglycans (CSPGs) and keratan sulfate proteoglycans (KSPGs) in directing mossy fiber (MF) outgrowth and regeneration in rat hippocampal slice cultures. MFs normally exhibit a very specific innervation pattern that is restricted to the stratum lucidum (SL). In addition, MFs in hippocampal slice cultures will regenerate this specific innervation pattern after transection. CSPGs are one of the best characterized inhibitory axon guidance molecules in the CNS and are widely expressed in all areas of the hippocampus except SL. KSPGs are also widely expressed in the hippocampus, but their role in axon outgrowth has not been extensively studied in the CNS where phosphacan is the only protein that appears to contain KS-GAGs. Cultured hippocampal slices were treated with either chondroitin ABC lyase or keratanases to reduce the inhibitory axon guidance properties of CS and KS proteoglycans, respectively. The ability of transected MFs to regenerate their normal innervation pattern after digestion of CS and KS-GAGS sugars with these enzymes was examined. Only keratanase treatment resulted in misrouting of MFs. Identifying the mechanism by which keratanase produced MF misrouting is complicated by the presence of splice variants of the phosphacan gene that include the extracellular form of phosphacan and the transmembrane receptor protein tyrosine phosphatase beta/zeta (RPTPbeta/zeta). Both forms of phosphacan are made by astrocytes, suggesting that keratanase alters MF outgrowth by modifying astrocyte function.
Figures


References
-
- Altman J, Das GD (1965) Autoradiographic and histological evidence of postnatal hippocampal neurogenesis in rats. J Comp Neurol 124: 319-336. - PubMed
-
- Alvarez-Royo P, Zola-Morgan S, Squire LR (1992) Impairment of long-term memory and sparing of short-term memory in monkeys with medial temporal lobe lesions: a response to Ringo. Behav Brain Res 52: 1-5. - PubMed
-
- Asher RA, Morgenstern DA, Moon LD, Fawcett JW (2001) Chondroitin sulphate proteoglycans: inhibitory components of the glial scar. Prog Brain Res 132: 611-619. - PubMed
-
- Brittis PA, Canning DR, Silver J (1992) Chondroitin sulfate as a regulator of neuronal patterning in the retina. Science 255: 733-736. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources