

Pathogenicity islands in bacterial pathogenesis
- PMID: 14726454
- PMCID: PMC321463
- DOI: 10.1128/CMR.17.1.14-56.2004
Pathogenicity islands in bacterial pathogenesis
Erratum in
- Clin Microbiol Rev. 2006 Jan;19(1):257
Abstract
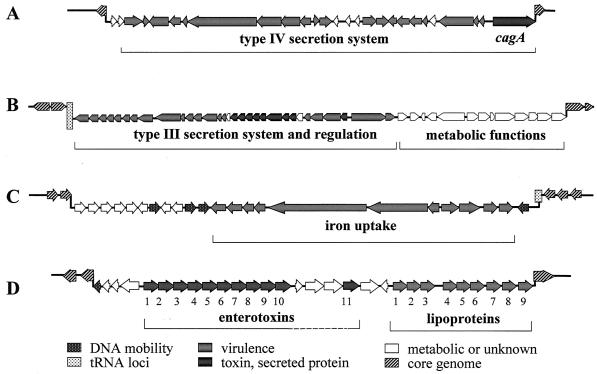
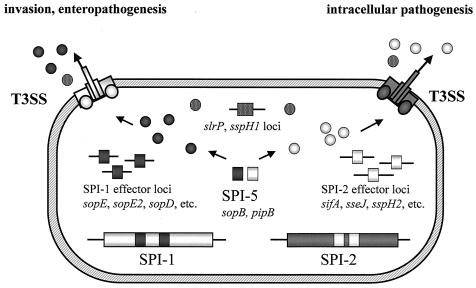
In this review, we focus on a group of mobile genetic elements designated pathogenicity islands (PAI). These elements play a pivotal role in the virulence of bacterial pathogens of humans and are also essential for virulence in pathogens of animals and plants. Characteristic molecular features of PAI of important human pathogens and their role in pathogenesis are described. The availability of a large number of genome sequences of pathogenic bacteria and their benign relatives currently offers a unique opportunity for the identification of novel pathogen-specific genomic islands. However, this knowledge has to be complemented by improved model systems for the analysis of virulence functions of bacterial pathogens. PAI apparently have been acquired during the speciation of pathogens from their nonpathogenic or environmental ancestors. The acquisition of PAI not only is an ancient evolutionary event that led to the appearance of bacterial pathogens on a timescale of millions of years but also may represent a mechanism that contributes to the appearance of new pathogens within a human life span. The acquisition of knowledge about PAI, their structure, their mobility, and the pathogenicity factors they encode not only is helpful in gaining a better understanding of bacterial evolution and interactions of pathogens with eukaryotic host cells but also may have important practical implications such as providing delivery systems for vaccination, tools for cell biology, and tools for the development of new strategies for therapy of bacterial infections.
Figures
References
-
- Akopyants, N. S., S. W. Clifton, D. Kersulyte, J. E. Crabtree, B. E. Youree, C. A. Reece, N. O. Bukanov, E. S. Drazek, B. A. Roe, and D. E. Berg. 1998. Analyses of the cag pathogenicity island of Helicobacter pylori. Mol. Microbiol. 28:37-53. - PubMed
-
- Alfano, J. R., A. O. Charkowski, W. L. Deng, J. L. Badel, T. Petnicki-Ocwieja, K. van Dijk, and A. Collmer. 2000. The Pseudomonas syringae Hrp pathogenicity island has a tripartite mosaic structure composed of a cluster of type III secretion genes bounded by exchangeable effector and conserved effector loci that contribute to parasitic fitness and pathogenicity in plants. Proc. Natl. Acad. Sci. USA 97:4856-4861. - PMC - PubMed
-
- Al-Hasani, K., I. R. Henderson, H. Sakellaris, K. Rajakumar, T. Grant, J. P. Nataro, R. Robins-Browne, and B. Adler. 2000. The sigA gene which is borne on the she pathogenicity island of Shigella flexneri 2a encodes an exported cytopathic protease involved in intestinal fluid accumulation. Infect. Immun. 68:2457-2463. - PMC - PubMed
-
- Allison, G. E., and N. K. Verma. 2000. Serotype-converting bacteriophages and O-antigen modification in Shigella flexneri. Trends Microbiol. 8:17-23. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
