

The Arabidopsis homologue of Xrcc3 plays an essential role in meiosis
- PMID: 14726957
- PMCID: PMC1271761
- DOI: 10.1038/sj.emboj.7600055
The Arabidopsis homologue of Xrcc3 plays an essential role in meiosis
Abstract
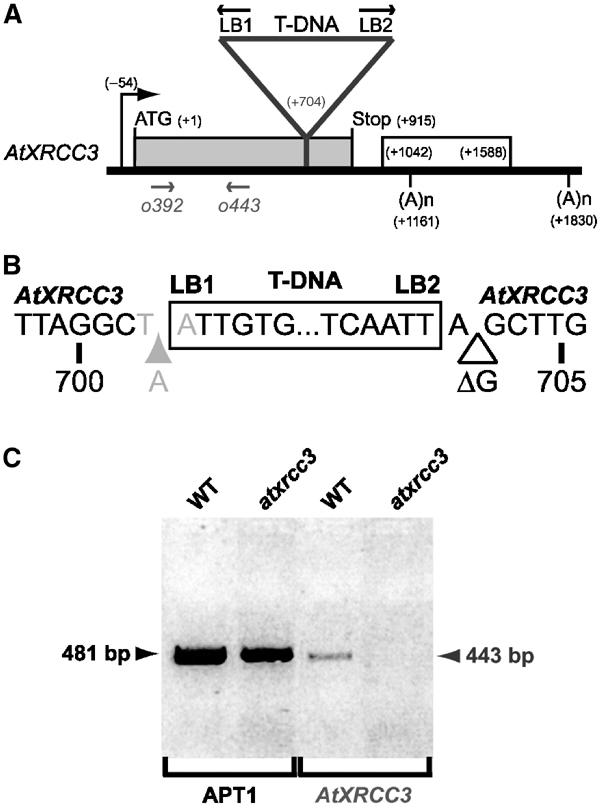
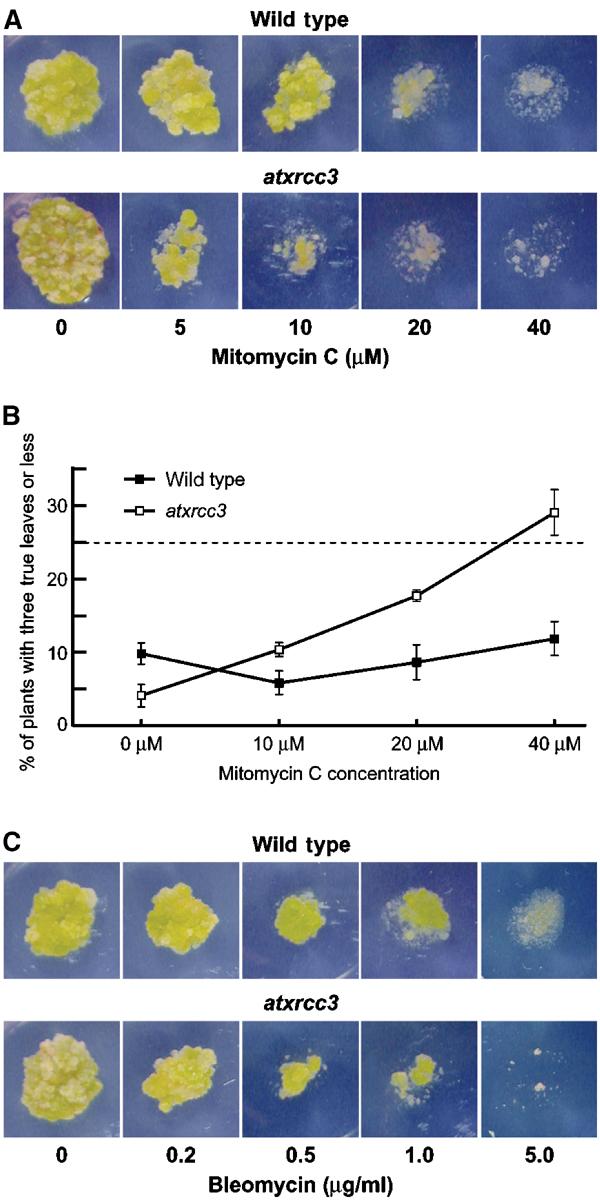
The eukaryotic RecA homologue Rad51 is a key factor in homologous recombination and recombinational repair. Rad51-like proteins have been identified from yeast (Rad55, Rad57 and Dmc1) to vertebrates (Rad51B, Rad51C, Rad51D, Xrcc2, Xrcc3 and Dmc1). These Rad51-like proteins are all members of the genetic recombination and DNA damage repair pathways. The sequenced genome of Arabidopsis thaliana encodes putative homologues of all six vertebrate Rad51-like proteins. We have identified and characterized an Arabidopsis mutant defective for one of these, AtXRCC3, the homologue of XRCC3. atxrcc3 plants are sterile, while they have normal vegetative development. Cytological observation shows that the atxrcc3 mutation does not affect homologous chromosome synapsis, but leads to chromosome fragmentation after pachytene, thus disrupting both male and female gametogenesis. This study shows an essential role for AtXrcc3 in meiosis in plants and possibly in other higher eukaryotes. Furthermore, atxrcc3 cells and plants are hypersensitive to DNA-damaging treatments, supporting the involvement of this Arabidopsis Rad51-like protein in recombinational repair.
Figures





References
-
- Alexander MP (1969) Differential staining of aborted and nonaborted pollen. Stain Technol 44: 117–122 - PubMed
-
- Alonso JM, Stepanova AN, Leisse TJ, Kim CJ, Chen H, Shinn P, Stevenson DK, Zimmerman J, Barajas P, Cheuk R, Gadrinab C, Heller C, Jeske A, Koesema E, Meyers CC, Parker H, Prednis L, Ansari Y, Choy N, Deen H, Geralt M, Hazari N, Hom E, Karnes M, Mulholland C, Ndubaku R, Schmidt I, Guzman P, Aguilar-Henonin L, Schmid M, Weigel D, Carter DE, Marchand T, Risseeuw E, Brogden D, Zeko A, Crosby WL, Berry CC, Ecker JR (2003) Genome-wide insertional mutagenesis of Arabidopsis thaliana. Science 301: 653–657 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
