

A plasma membrane protein from Zea mays binds with the herbivore elicitor volicitin
- PMID: 14729912
- PMCID: PMC341921
- DOI: 10.1105/tpc.017723
A plasma membrane protein from Zea mays binds with the herbivore elicitor volicitin
Abstract
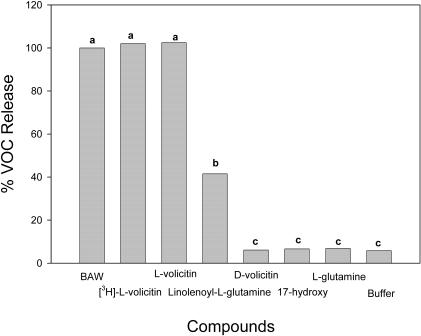
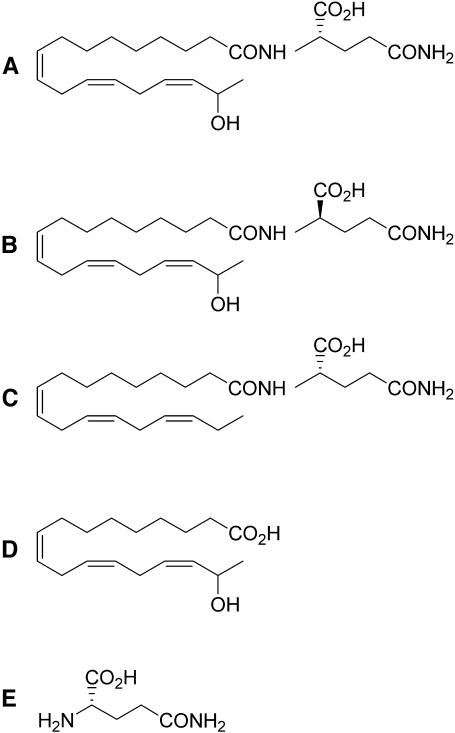
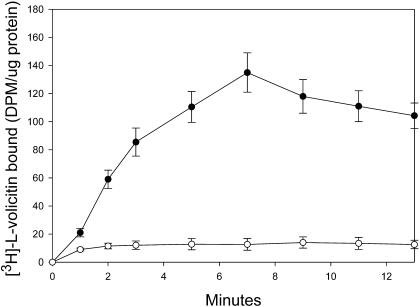
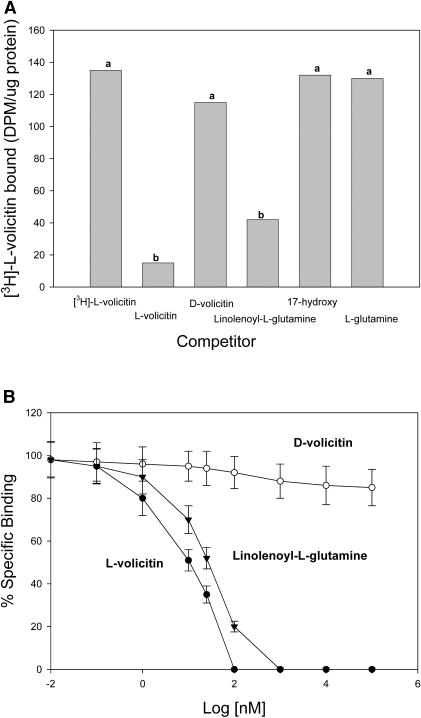
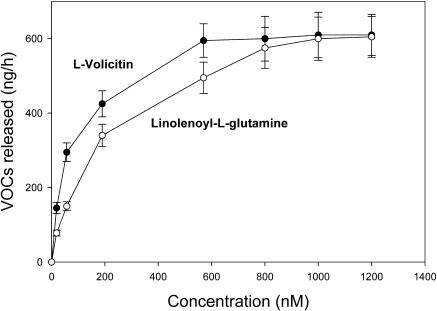
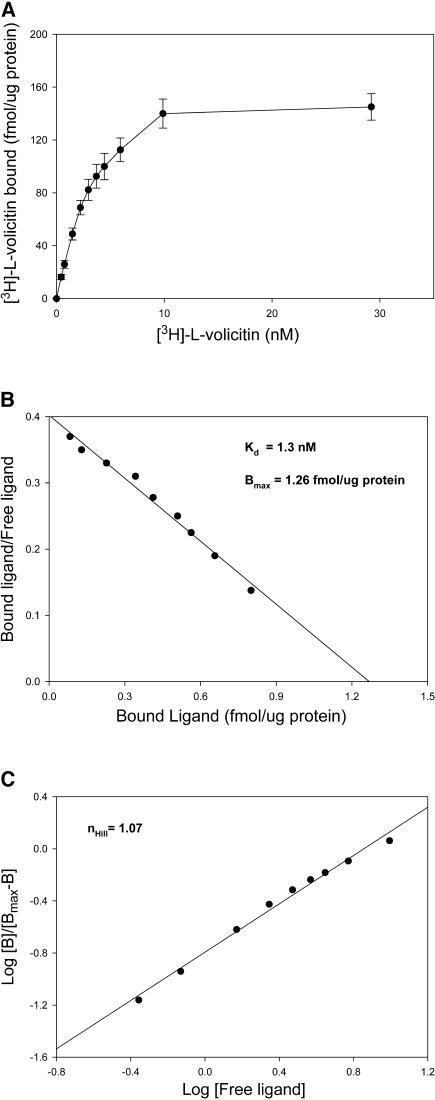
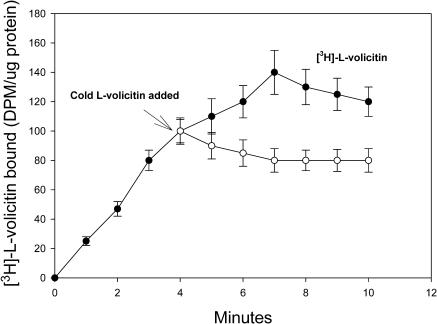
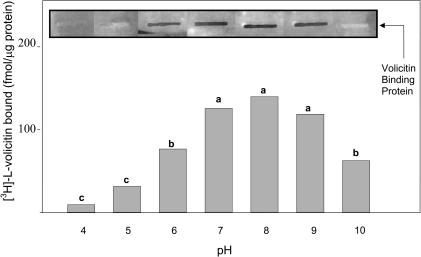
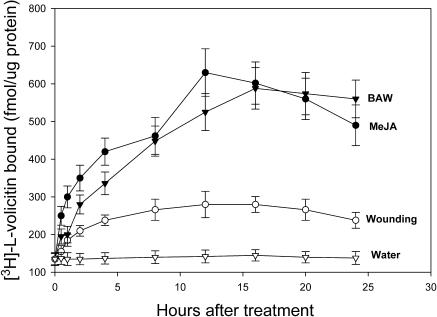
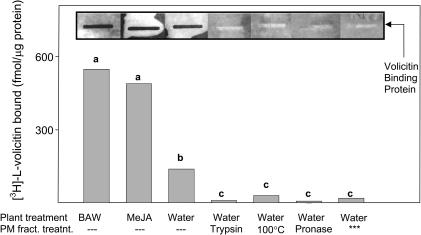
Volicitin (17-hydroxylinolenoyl-l-Gln) present in the regurgitant of Spodoptera exigua (beet armyworm caterpillars) activates the emission of volatile organic compounds (VOCs) when in contact with damaged Zea mays cv Delprim (maize) leaves. VOC emissions in turn serve as a signaling defense for the plant by attracting female parasitic wasps that prey on herbivore larvae. A tritiated form of volicitin was synthesized and shown to induce volatiles in the same fashion as the biological form. [(3)H]-l-volicitin rapidly, reversibly, and saturably bound to enriched plasma membrane fractions isolated from Z. mays leaves with an apparent K(d) of 1.3 nM and a Hill coefficient of 1.07. Analog studies showed that the l-Gln and hydroxy moieties of volicitin play an important role in binding. Treatment of plants with methyl jasmonate (MeJA) increased the total binding of [(3)H]-l-volicitin to the enriched plasma membrane more than threefold, suggesting that MeJA activates transcription of the gene encoding the binding protein. S. exigua feeding also increased total binding fourfold. Cycloheximide pretreatment of plants significantly decreased binding of radiolabeled volicitin to the enriched plasma membrane. These data provide the first experimental evidence that initiation of plant defenses in response to herbivore damage can be mediated by a binding protein-ligand interaction.
Figures
Similar articles
-
In situ translocation of volicitin by beet armyworm larvae to maize and systemic immobility of the herbivore elicitor in planta.Planta. 2004 Apr;218(6):999-1007. doi: 10.1007/s00425-003-1173-6. Epub 2003 Dec 18. Planta. 2004. PMID: 14685859
-
The influence of intact-plant and excised-leaf bioassay designs on volicitin- and jasmonic acid-induced sesquiterpene volatile release in Zea mays.Planta. 2001 Dec;214(2):171-9. doi: 10.1007/s004250100603. Planta. 2001. PMID: 11800380
-
Maize genes induced by herbivory and volicitin.J Chem Ecol. 2004 Dec;30(12):2543-57. doi: 10.1007/s10886-004-7949-8. J Chem Ecol. 2004. PMID: 15724970
-
Elicitors and priming agents initiate plant defense responses.Photosynth Res. 2005 Aug;85(2):149-59. doi: 10.1007/s11120-005-1001-x. Photosynth Res. 2005. PMID: 16075316 Review.
-
[Herbivore elicitors and plant signaling pathways involved in the production of herbivore-induced plant volatiles].Tanpakushitsu Kakusan Koso. 2003 Oct;48(13):1786-92. Tanpakushitsu Kakusan Koso. 2003. PMID: 14535205 Review. Japanese. No abstract available.
Cited by
-
Caterpillar-Induced Volatile Emissions in Cotton: The Relative Importance of Damage and Insect-Derived Factors.Front Plant Sci. 2021 Aug 3;12:709858. doi: 10.3389/fpls.2021.709858. eCollection 2021. Front Plant Sci. 2021. PMID: 34413869 Free PMC article.
-
Cowpea chloroplastic ATP synthase is the source of multiple plant defense elicitors during insect herbivory.Plant Physiol. 2007 Jun;144(2):793-805. doi: 10.1104/pp.107.097154. Epub 2007 Mar 16. Plant Physiol. 2007. PMID: 17369425 Free PMC article.
-
The Biosynthesis and Metabolism of the N-Acylated Aromatic Amino Acids: N-Acylphenylalanine, N-Acyltyrosine, N-Acyltryptophan, and N-Acylhistidine.Front Mol Biosci. 2022 Jan 3;8:801749. doi: 10.3389/fmolb.2021.801749. eCollection 2021. Front Mol Biosci. 2022. PMID: 35047560 Free PMC article. Review.
-
Do caterpillars secrete "oral secretions"?J Chem Ecol. 2009 Mar;35(3):326-35. doi: 10.1007/s10886-009-9604-x. Epub 2009 Feb 17. J Chem Ecol. 2009. PMID: 19221841
-
Infestation by the Piercing-Sucking Herbivore Nilaparvata lugens Systemically Triggers JA- and SA-Dependent Defense Responses in Rice.Biology (Basel). 2023 Jun 5;12(6):820. doi: 10.3390/biology12060820. Biology (Basel). 2023. PMID: 37372105 Free PMC article.
References
-
- Alborn, H.T., Turlings, T.C.J., Jones, T.H., Stenhagen, G., Loughrin, J.H., and Tumlinson, J.H. (1997). An elicitor of plant volatiles from beet armyworm oral secretion. Science 276, 945–949.
-
- Ames, B.N. (1966). Assay of inorganic phosphate, total phosphate and phosphatase. Meth. Enzymol. 8, 115–118.
-
- Blee, E. (2002). Impact of phyto-oxylipins in plant defense. Trends Plant Sci. 7, 315–321. - PubMed
-
- Bradford, M.M. (1976). A rapid and sensitive method for the quantification of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 72, 248–255. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
