

Drosophila melanogaster MNK/Chk2 and p53 regulate multiple DNA repair and apoptotic pathways following DNA damage
- PMID: 14729967
- PMCID: PMC321428
- DOI: 10.1128/MCB.24.3.1219-1231.2004
Drosophila melanogaster MNK/Chk2 and p53 regulate multiple DNA repair and apoptotic pathways following DNA damage
Abstract
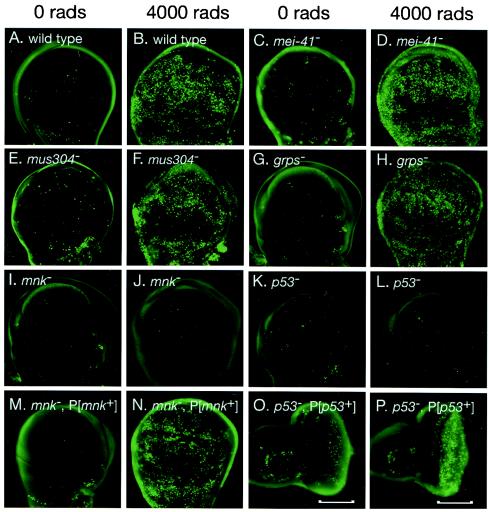
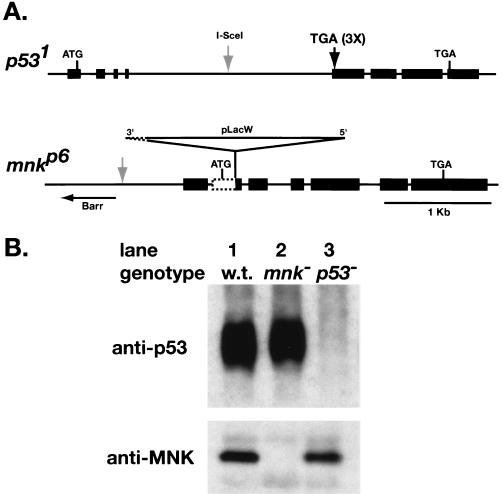
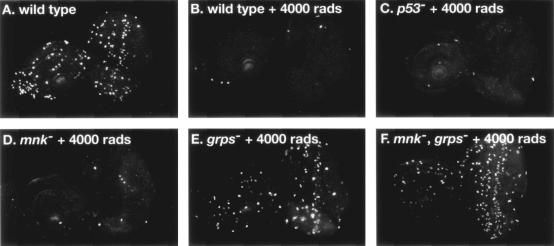
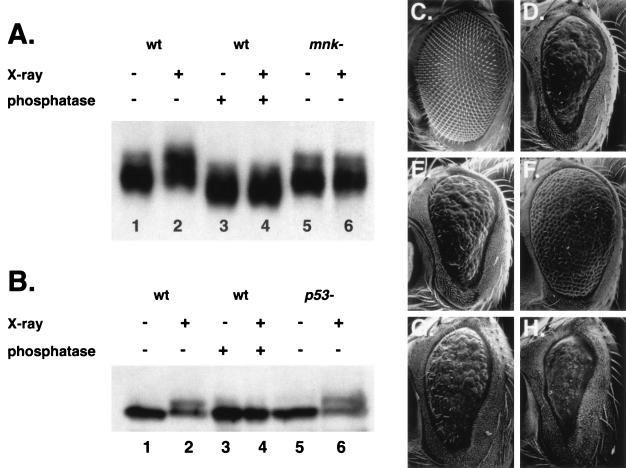
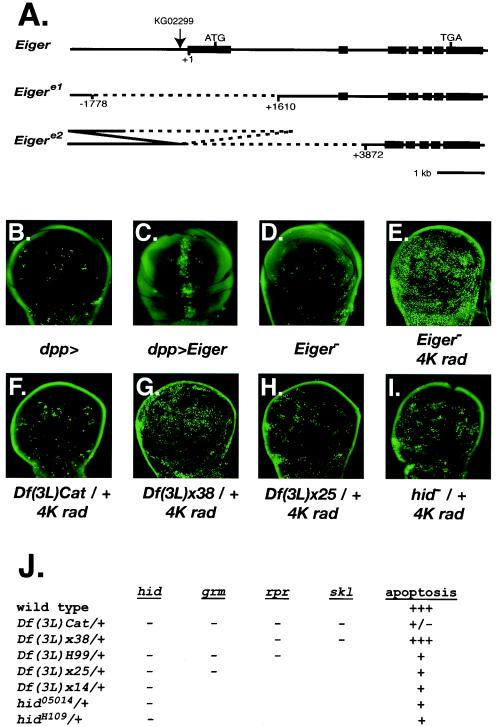
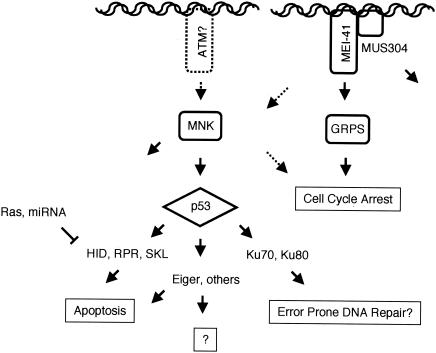
We have used genetic and microarray analysis to determine how ionizing radiation (IR) induces p53-dependent transcription and apoptosis in Drosophila melanogaster. IR induces MNK/Chk2-dependent phosphorylation of p53 without changing p53 protein levels, indicating that p53 activity can be regulated without an Mdm2-like activity. In a genome-wide analysis of IR-induced transcription in wild-type and mutant embryos, all IR-induced increases in transcript levels required both p53 and the Drosophila Chk2 homolog MNK. Proapoptotic targets of p53 include hid, reaper, sickle, and the tumor necrosis factor family member EIGER: Overexpression of Eiger is sufficient to induce apoptosis, but mutations in Eiger do not block IR-induced apoptosis. Animals heterozygous for deletions that span the reaper, sickle, and hid genes exhibited reduced IR-dependent apoptosis, indicating that this gene complex is haploinsufficient for induction of apoptosis. Among the genes in this region, hid plays a central, dosage-sensitive role in IR-induced apoptosis. p53 and MNK/Chk2 also regulate DNA repair genes, including two components of the nonhomologous end-joining repair pathway, Ku70 and Ku80. Our results indicate that MNK/Chk2-dependent modification of Drosophila p53 activates a global transcriptional response to DNA damage that induces error-prone DNA repair as well as intrinsic and extrinsic apoptosis pathways.
Figures
References
-
- Abdu, U., M. Brodsky, and T. Schupbach. 2002. Activation of a meiotic checkpoint during Drosophila oogenesis regulates the translation of Gurken through Chk2/Mnk. Curr. Biol. 12:1645. - PubMed
-
- Abraham, R. T. 2001. Cell cycle checkpoint signaling through the ATM and ATR kinases. Genes Dev. 15:2177-2196. - PubMed
-
- Abrams, J. M., K. White, L. I. Fessler, and H. Steller. 1993. Programmed cell death during Drosophila embryogenesis. Development 117:29-43. - PubMed
-
- Amundson, S. A., M. Bittner, P. Meltzer, J. Trent, and A. J. Fornace, Jr. 2001. Physiological function as regulation of large transcriptional programs: the cellular response to genotoxic stress. Comp. Biochem. Physiol. B 129:703-710. - PubMed
-
- Barlow, C., K. D. Brown, C. X. Deng, D. A. Tagle, and A. Wynshaw-Boris. 1997. Atm selectively regulates distinct p53-dependent cell-cycle checkpoint and apoptotic pathways. Nat. Genet. 17:453-456. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous