

RNA folding affects the recruitment of SR proteins by mouse and human polypurinic enhancer elements in the fibronectin EDA exon
- PMID: 14729981
- PMCID: PMC321440
- DOI: 10.1128/MCB.24.3.1387-1400.2004
RNA folding affects the recruitment of SR proteins by mouse and human polypurinic enhancer elements in the fibronectin EDA exon
Abstract
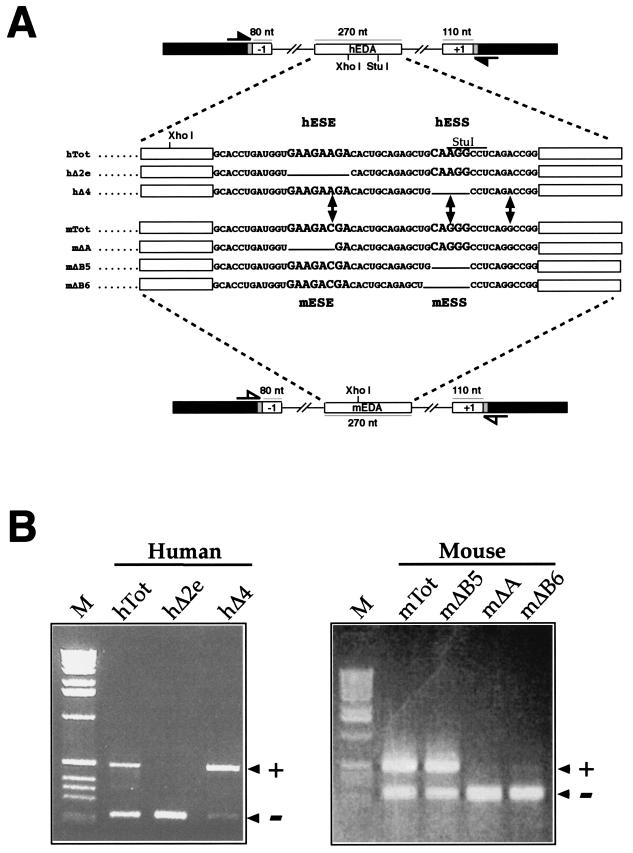
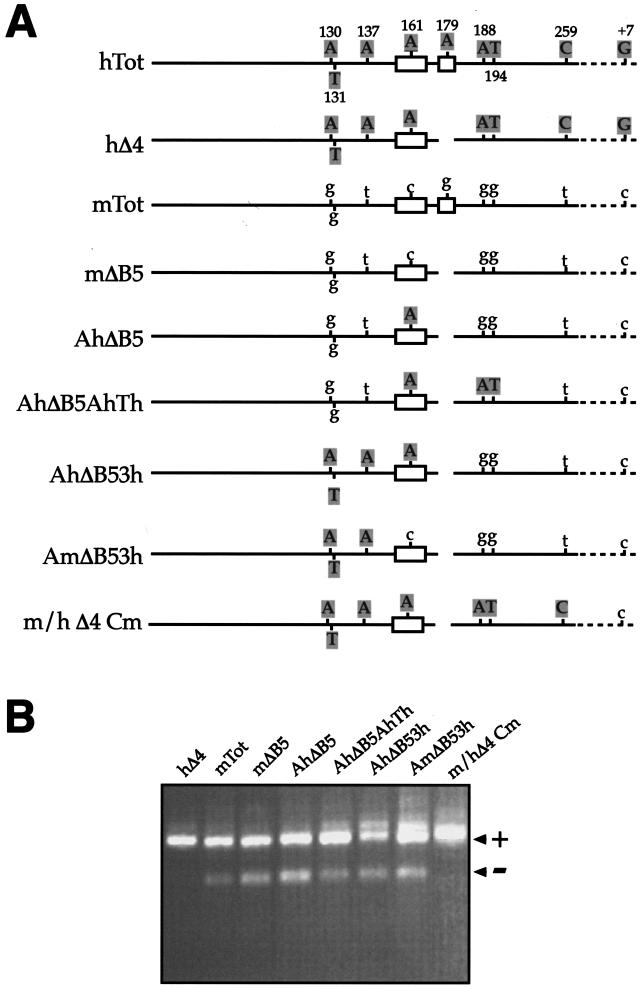
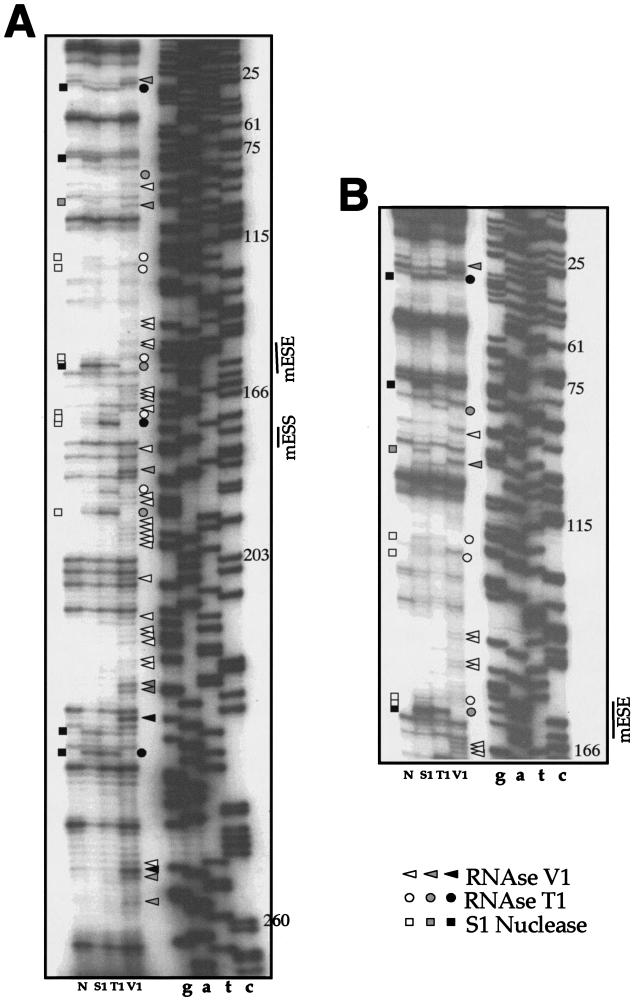
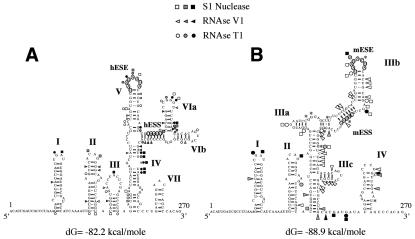
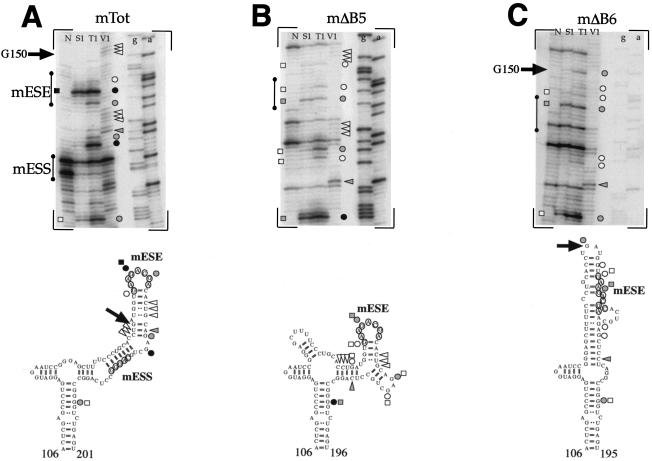
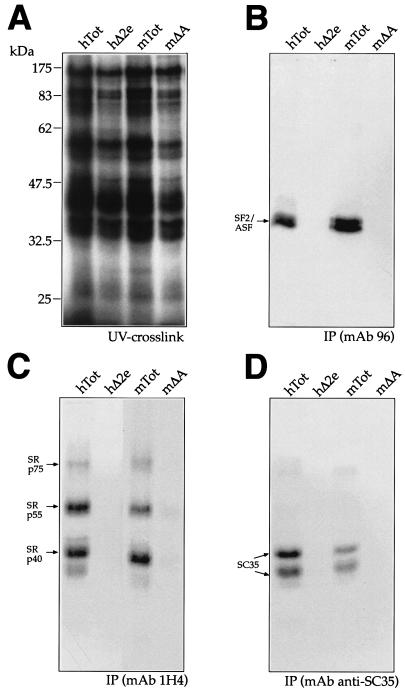
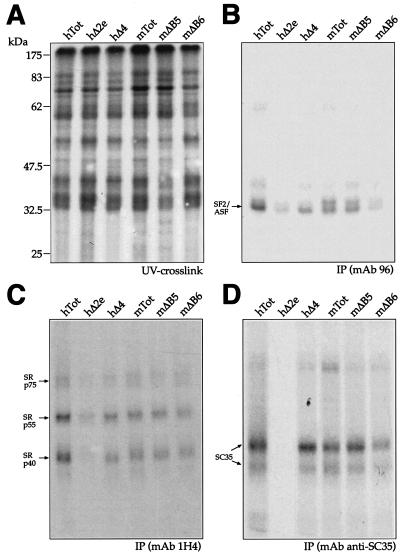
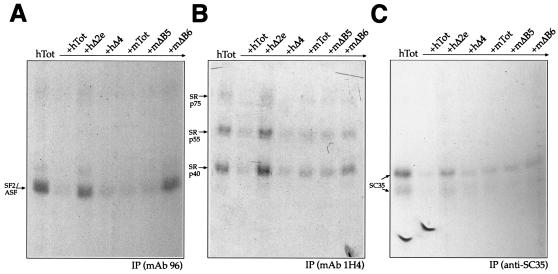
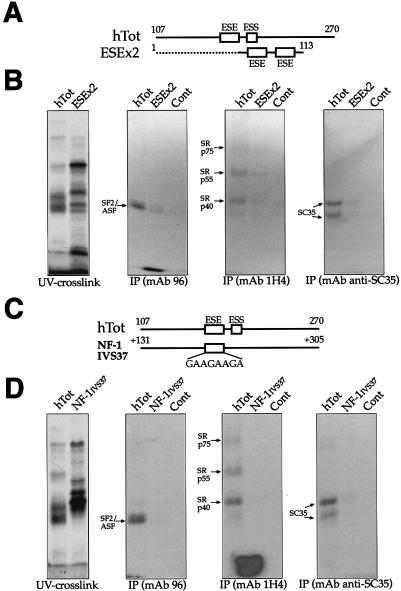
In humans, inclusion or exclusion of the fibronectin EDA exon is mainly regulated by a polypurinic enhancer element (exonic splicing enhancer [ESE]) and a nearby silencer element (exonic splicing silencer [ESS]). While human and mouse ESEs behave identically, mutations introduced into the homologous mouse ESS sequence result either in no change in splicing efficiency or in complete exclusion of the exon. Here, we show that this apparently contradictory behavior cannot be simply accounted for by a localized sequence variation between the two species. Rather, the nucleotide differences as a whole determine several changes in the respective RNA secondary structures. By comparing how the two different structures respond to homologous deletions in their putative ESS sequences, we show that changes in splicing behavior can be accounted for by a differential ESE display in the two RNAs. This is confirmed by RNA-protein interaction analysis of levels of SR protein binding to each exon. The immunoprecipitation patterns show the presence of complex multi-SR protein-RNA interactions that are lost with secondary-structure variations after the introduction of ESE and ESS variations. Taken together, our results demonstrate that the sequence context, in addition to the primary sequence identity, can heavily contribute to the making of functional units capable of influencing pre-mRNA splicing.
Figures
References
-
- Adams, M. D., D. Z. Rudner, and D. C. Rio. 1996. Biochemistry and regulation of pre-mRNA splicing. Curr. Opin. Cell Biol. 8:331-339. - PubMed
-
- Balvay, L., D. Libri, and M. Y. Fiszman. 1993. Pre-mRNA secondary structure and the regulation of splicing. Bioessays 15:165-169. - PubMed
-
- Black, D. L. 1991. Does steric interference between splice sites block the splicing of a short c-src neuron-specific exon in non-neuronal cells? Genes Dev. 5:389-402. - PubMed
-
- Blencowe, B. J. 2000. Exonic splicing enhancers: mechanism of action, diversity and role in human genetic diseases. Trends Biochem. Sci. 25:106-110. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials