

Vertebrate ultraviolet visual pigments: protonation of the retinylidene Schiff base and a counterion switch during photoactivation
- PMID: 14732701
- PMCID: PMC327121
- DOI: 10.1073/pnas.0305206101
Vertebrate ultraviolet visual pigments: protonation of the retinylidene Schiff base and a counterion switch during photoactivation
Abstract
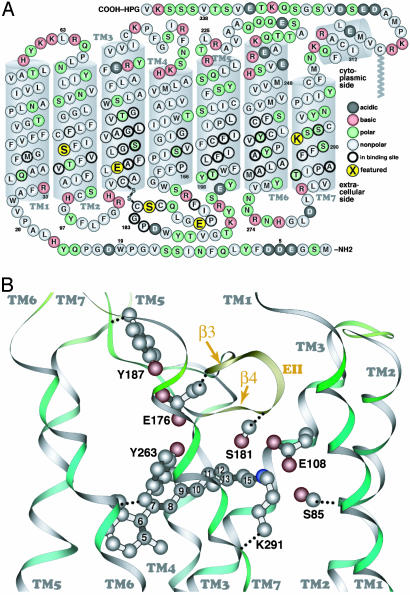
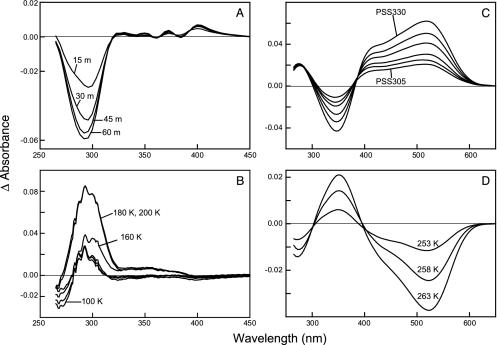
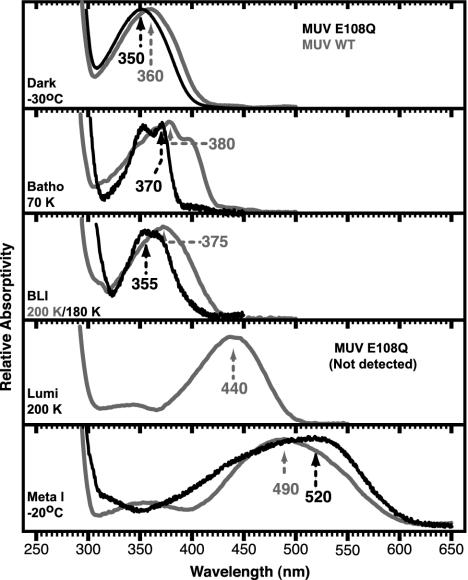
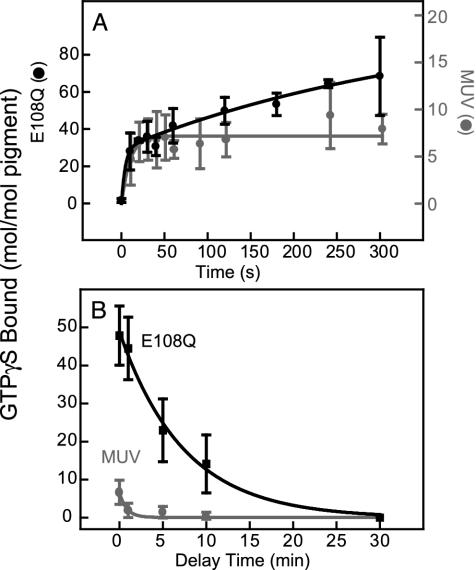
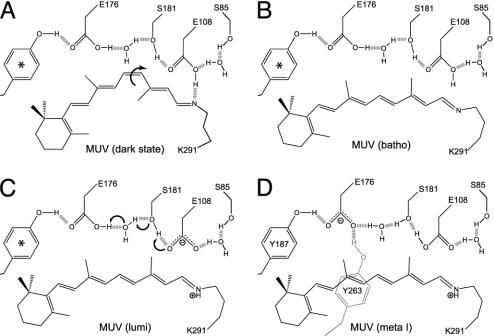
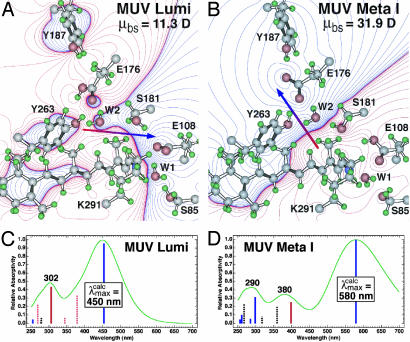
For visual pigments, a covalent bond between the ligand (11-cis-retinal) and receptor (opsin) is crucial to spectral tuning and photoactivation. All photoreceptors have retinal bound via a Schiff base (SB) linkage, but only UV-sensitive cone pigments have this moiety unprotonated in the dark. We investigated the dynamics of mouse UV (MUV) photoactivation, focusing on SB protonation and the functional role of a highly conserved acidic residue (E108) in the third transmembrane helix. On illumination, wild-type MUV undergoes a series of conformational changes, batho --> lumi --> meta I, finally forming the active intermediate meta II. During the dark reactions, the SB becomes protonated transiently. In contrast, the MUV-E108Q mutant formed significantly less batho that did not decay through a protonated lumi. Rather, a transition to meta I occurred above approximately 240 K, with a remarkable red shift (lambda(max) approximately 520 nm) accompanying SB protonation. The MUV-E108Q meta I --> meta II transition appeared normal but the MUV-E108Q meta II decay to opsin and free retinal was dramatically delayed, resulting in increased transducin activation. These results suggest that there are two proton donors during the activation of UV pigments, the primary counterion E108 necessary for protonation of the SB during lumi formation and a second one necessary for protonation of meta I. Inactivation of meta II in SWS1 cone pigments is regulated by the primary counterion. Computational studies suggest that UV pigments adopt a switch to a more distant counterion, E176, during the lumi to meta I transition. The findings with MUV are in close analogy to rhodopsin and provides further support for the importance of the counterion switch in the photoactivation of both rod and cone visual pigments.
Figures
References
-
- Ebrey, T. (2000) Methods Enzymol. 315, 196–207. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
