

The distributions of mitochondria and sodium channels reflect the specific energy requirements and conduction properties of the human optic nerve head
- PMID: 14736793
- PMCID: PMC1771975
- DOI: 10.1136/bjo.2003.027664
The distributions of mitochondria and sodium channels reflect the specific energy requirements and conduction properties of the human optic nerve head
Abstract
Aim: To study the normal distributions of mitochondria and voltage gated Na+ channels in the human optic nerve head in order to gain insight into the potential mechanisms of optic nerve dysfunction seen in the inherited optic neuropathies.
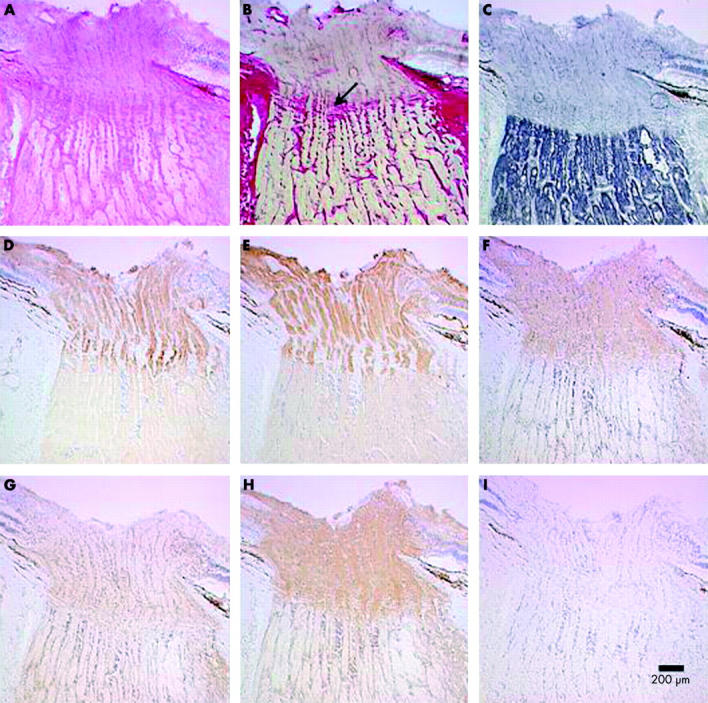
Methods: Five fresh frozen human optic nerves were studied. Longitudinally orientated, serial cryosections of optic nerve head were cut for mitochondrial enzyme histochemistry and immunolabelling for cytochrome c oxidase (COX) subunits and voltage gated Na+ channel subtypes (Na(v) 1.1, 1.2, 1.3, and 1.6).
Results: A high density of voltage gated Na+ channels (subtypes Na(v) 1.1, 1.3, and 1.6) in the unmyelinated, prelaminar, and laminar optic nerve was found. This distribution co-localised both with areas of high COX activity and strong immunolabelling for COX subunits I and IV.
Conclusions: Increased numbers of mitochondria in the prelaminar optic nerve have previously been interpreted as indicating a mechanical hold up of axoplasmic flow at the lamina cribrosa. These results suggest that this increased mitochondrial density serves the higher energy requirements for electrical conduction in unmyelinated axons in the prelaminar and laminar optic nerve and is not a reflection of any mechanical restriction. This could explain why optic neuropathies typically occur in primary inherited mitochondrial diseases such as Leber's hereditary optic neuropathy, myoclonic epilepsy with ragged red fibres (MERRF), and Leigh's syndrome. Secondary mitochondrial dysfunction has also been reported in dominant optic atrophy, Friedreich's ataxia, tobacco alcohol amblyopia, Cuban epidemic optic neuropathy, and chloramphenicol optic neuropathy. These diseases are rare but these findings challenge the traditional theories of optic nerve structure and function and may suggest an alternative approach to the study of commoner optic neuropathies such as glaucoma.
Figures

References
-
- Minckler DS, McLean IW, Tso MO. Distribution of axonal and glial elements in the rhesus optic nerve head studied by electron microscopy. Am J Ophthalmol 1976;82:179–87. - PubMed
-
- Hollander H, Makarov F, Stefani FH, et al. Evidence of constriction of optic nerve axons at the lamina cribrosa in the normotensive eye in humans and other mammals. Ophthalmic Res 1995;27:296–309. - PubMed
-
- Bristow EA, Griffiths PG, Andrews RM, et al. The distribution of mitochondrial activity in relation to optic nerve structure. Arch Ophthalmol 2002;120:791–6. - PubMed
-
- Bancroft JD, Stevens A. Theory and practice of histological techniques. Edinburgh: Churchill Livingstone, 1996.
MeSH terms
Substances
LinkOut - more resources
Full Text Sources