

Calcium signaling is involved in dynein-dependent microtubule organization
- PMID: 14742707
- PMCID: PMC379291
- DOI: 10.1091/mbc.e03-09-0675
Calcium signaling is involved in dynein-dependent microtubule organization
Abstract
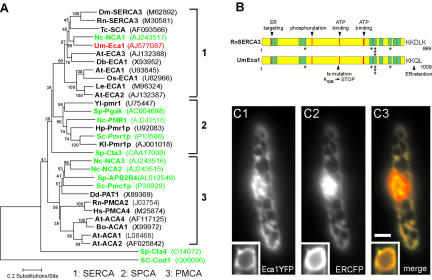
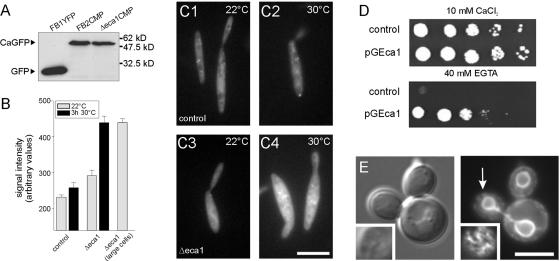
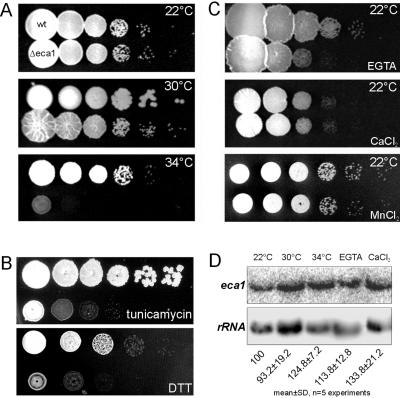
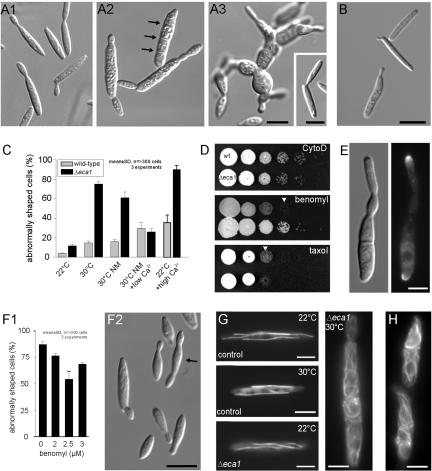
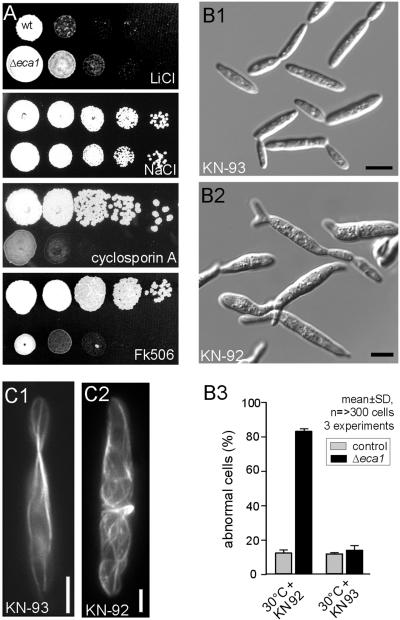
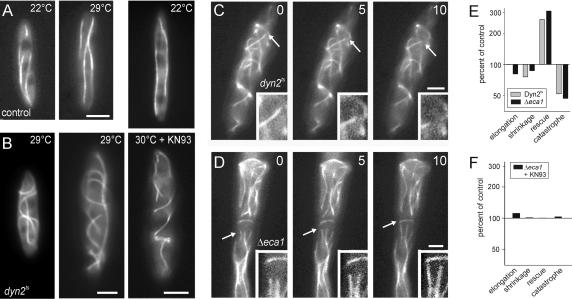
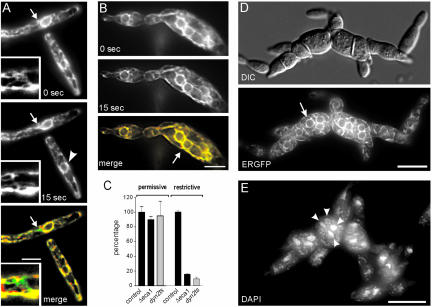
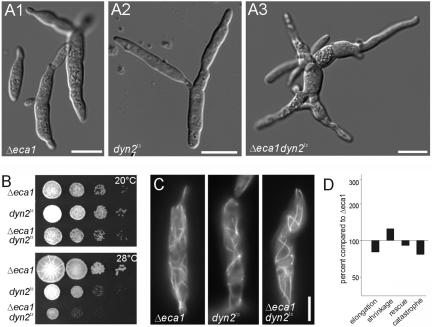
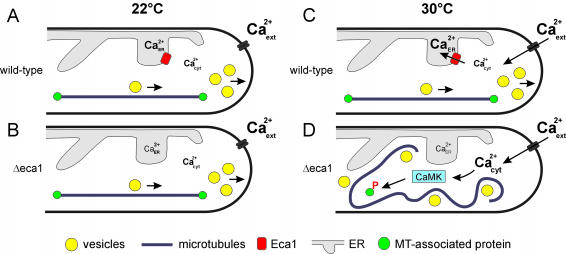
The microtubule cytoskeleton supports cellular morphogenesis and polar growth, but the underlying mechanisms are not understood. In a screen for morphology mutants defective in microtubule organization in the fungus Ustilago maydis, we identified eca1 that encodes a sarcoplasmic/endoplasmic calcium ATPase. Eca1 resides in the endoplasmic reticulum and restores growth of a yeast mutant defective in calcium homeostasis. Deletion of eca1 resulted in elevated cytosolic calcium levels and a severe growth and morphology defect. While F-actin and myosin V distribution is unaffected, Deltaeca1 mutants contain longer and disorganized microtubules that show increased rescue and reduced catastrophe frequencies. Morphology can be restored by inhibition of Ca(2+)/calmodulin-dependent kinases or destabilizing microtubules, indicating that calcium-dependent alterations in dynamic instability are a major cause of the growth defect. Interestingly, dynein mutants show virtually identical changes in microtubule dynamics and dynein-dependent ER motility was drastically decreased in Deltaeca1. This indicates a connection between calcium signaling, dynein, and microtubule organization in morphogenesis of U. maydis.
Figures
References
-
- Andersson, H., Kappeler, F., and Hauri, H.P. (1999). Protein targeting to endoplasmic reticulum by dilysine signals involves direct retention in addition to retrieval. J. Biol. Chem. 274, 15080-15084. - PubMed
-
- Benito, B., Garciadeblas, B., and Rodriguez-Navarro, A. (2000). Molecular cloning of the calcium and sodium ATPases in Neurospora crassa. Mol. Microbiol. 35, 1079-1088. - PubMed
-
- Bölker, M. (2001). Ustilago maydis - a valuable model system for the study of fungal dimorphism and virulence. Microbiology 147, 1395-1401. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
