

Serotonergic facilitation of synaptic activity in the developing rat prefrontal cortex
- PMID: 14742723
- PMCID: PMC1665004
- DOI: 10.1113/jphysiol.2003.051284
Serotonergic facilitation of synaptic activity in the developing rat prefrontal cortex
Abstract
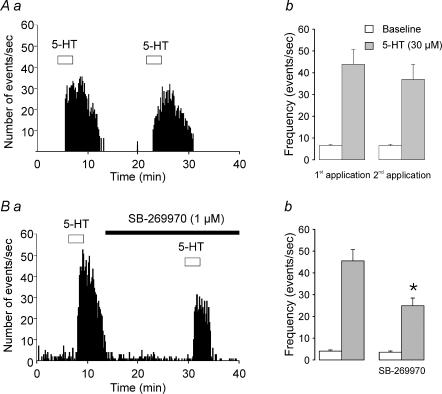
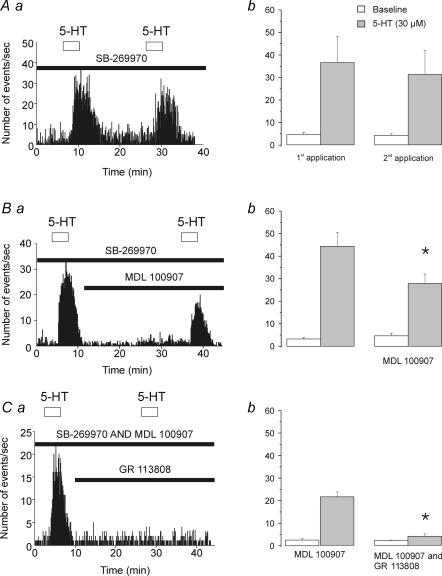
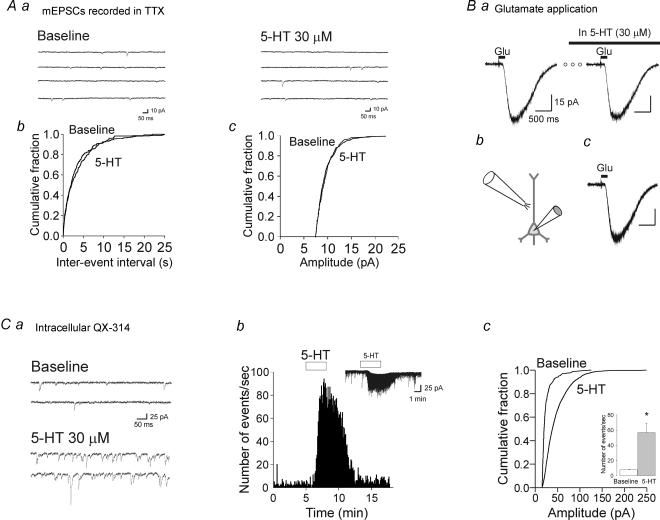
Previous studies have outlined an important role for serotonin (5-HT) in the development of synaptic connectivity and function in the cerebral cortex. In this study, we have examined the effects of 5-HT on synaptic function in prefrontal cortex at a time of intense synapse formation and remodelling. Whole-cell recordings in slices derived from animals aged postnatal (P) days 16-20 showed that administration of 5-HT induced a robust increase in synaptic activity that was blocked by CNQX but not by bicuculline. This 5-HT-induced increase in glutamate-mediated synaptic activity was pharmacologically heterogeneous as it was differentially inhibited by the receptor subtype-selective antagonists SB-269970, MDL 100907 and GR 113808 and thus involved 5-HT(7), 5-HT(2A) and 5-HT(4) receptors. These results, obtained in juvenile cortex, contrast with those seen in adults where the increase in spontaneous excitatory postsynaptic currents (sEPSCs) was mediated solely by 5-HT(2A) receptors. In developing cortex, activation of 5-HT(7), but not 5-HT(2A) or 5-HT(4) receptors, elicited a robust inward current. However, the facilitation of synaptic activity mediated by all three of these receptors involved increases in both the amplitude and frequency of sEPSCs and was blocked by TTX. These results are best interpreted as indicating that all three receptor subtypes increase synaptic activity by exciting neuronal elements within the slice. No evidence was found for a postsynaptic facilitation of synaptic currents by 5-HT. Together, these results show that the repertoire of electrophysiologically active 5-HT receptors in prefrontal cortex is developmentally regulated, and that 5-HT(7) and 5-HT(4) receptors play a previously unsuspected role in regulating synaptic activity in this region.
Figures





References
-
- Aghajanian GK, Marek GJ. Serotonin induces excitatory postsynaptic potentials in apical dendrites of neocortical pyramidal cells. Neuropharmacology. 1997;36:589–599. - PubMed
-
- Andrade R, Chaput Y. 5-Hydroxytryptamine4-like receptors mediate the slow excitatory response to serotonin in the rat hippocampus. J Pharmacol Exp Ther. 1991;257:930–937. - PubMed
-
- Araneda R, Andrade R. 5-Hydroxytryptamine2 and 5-hydroxytryptamine1A receptors mediate opposing responses on membrane excitability in rat association cortex. Neuroscience. 1991;40:399–412. - PubMed
-
- Bennett-Clarke CA, Lane RD, Rhoades RW. Fenfluramine depletes serotonin from the developing cortex and alters thalamocortical organization. Brain Res. 1995;702:255–260. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources