

Convergent evolution of disease resistance gene specificity in two flowering plant families
- PMID: 14742871
- PMCID: PMC341905
- DOI: 10.1105/tpc.016725
Convergent evolution of disease resistance gene specificity in two flowering plant families
Abstract
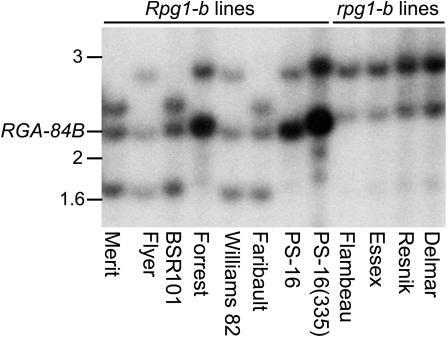
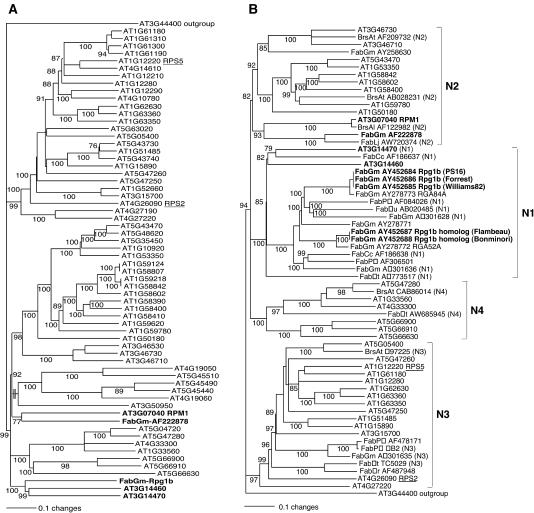
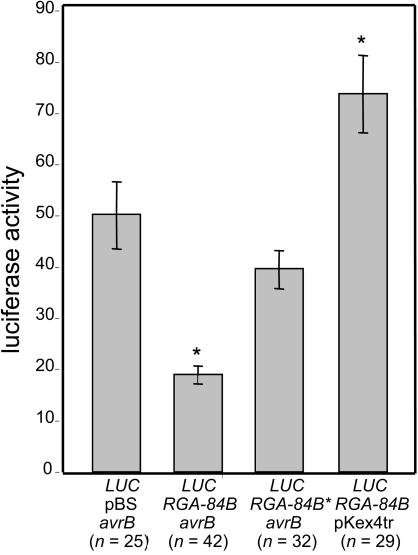
Plant disease resistance (R) genes that mediate recognition of the same pathogen determinant sometimes can be found in distantly related plant families. This observation implies that some R gene alleles may have been conserved throughout the diversification of land plants. To address this question, we have compared R genes from Glycine max (soybean), Rpg1-b, and Arabidopsis thaliana, RPM1, that mediate recognition of the same type III effector protein from Pseudomonas syringae, AvrB. RPM1 has been cloned previously, and here, we describe the isolation of Rpg1-b. Although RPM1 and Rpg1-b both belong to the coiled-coil nucleotide binding site (NBS) Leu-rich repeat (LRR) class of R genes, they share only limited sequence similarity outside the conserved domains characteristic of this class. Phylogenetic analyses of A. thaliana and legume NBS-LRR sequences demonstrate that Rpg1-b and RPM1 are not orthologous. We conclude that convergent evolution, rather than the conservation of an ancient specificity, is responsible for the generation of these AvrB-specific genes.
Figures



References
-
- Ashfield, T., Bocian, A., Held, D., Henk, A.D., Marek, L.F., Danesh, D., Peñuela, S., Meksem, K., Lightfoot, D.A., Young, N.D., Shoemaker, R.C., and Innes, R.W. (2003). Genetic and physical localization of the soybean Rpg1-b disease resistance gene reveals a complex locus containing several tightly linked families of NBS-LRR genes. Mol. Plant Microbe Interact. 16, 817–826. - PubMed
-
- Ashfield, T., Danzer, J.R., Held, D., Clayton, K., Keim, P., Saghai Maroof, M.A., Webb, P.M., and Innes, R.W. (1998). Rpg1, a soybean gene effective against races of bacterial blight, maps to a cluster of previously identified disease resistance genes. Theor. Appl. Genet. 96, 1013–1021.
-
- Axtell, M.J., and Staskawicz, B.J. (2003). Initiation of RPS2-specified disease resistance in Arabidopsis is coupled to the AvrRpt2-directed elimination of RIN4. Cell 112, 369–377. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
