

ELG1, a regulator of genome stability, has a role in telomere length regulation and in silencing
- PMID: 14745004
- PMCID: PMC341813
- DOI: 10.1073/pnas.0307796100
ELG1, a regulator of genome stability, has a role in telomere length regulation and in silencing
Abstract
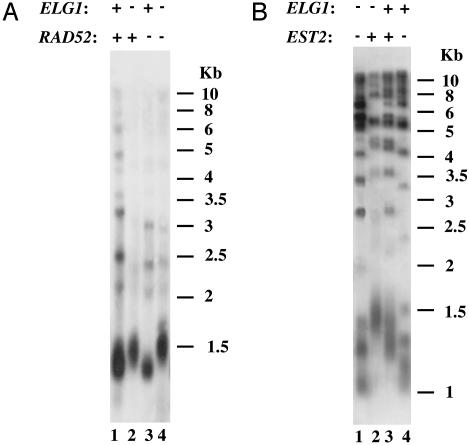
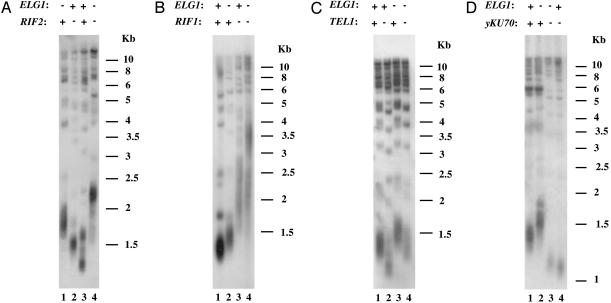
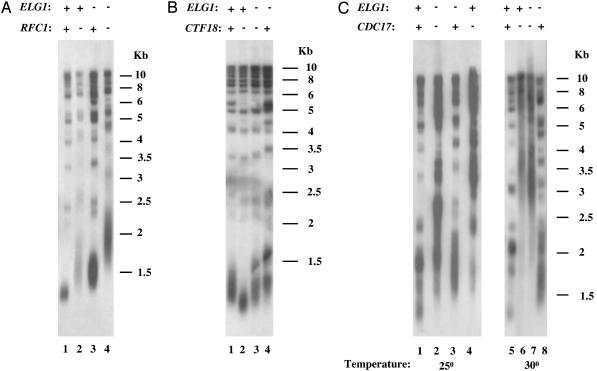
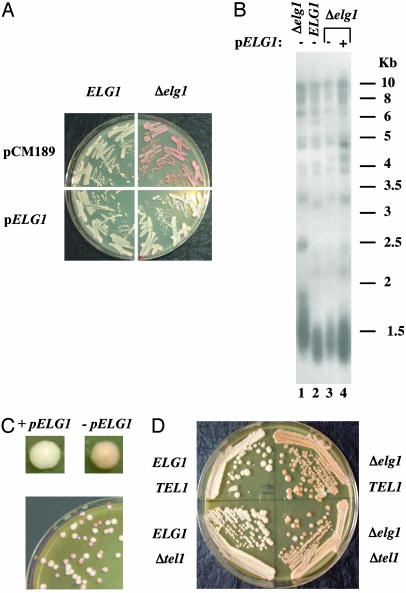
Telomeres, the natural ends of eukaryotic chromosomes, prevent the loss of chromosomal sequences and preclude their recognition as broken DNA. Telomere length is kept under strict boundaries by the action of various proteins, some with negative and others with positive effects on telomere length. Recently, data have been accumulating to support a role for DNA replication in the control of telomere length, although through a currently poorly understood mechanism. Elg1p, a replication factor C (RFC)-like protein of yeast, contributes to genome stability through a putative replication-associated function. Here, we show that Elg1p participates in negative control of telomere length and in telomeric silencing through a replication-mediated pathway. We show that the telomeric function of Elg1 is independent of recombination and completely dependent on an active telomerase. Additionally, this function depends on yKu and DNA polymerase. We discuss alternative models to explain how Elg1p affects telomere length.
Figures
References
-
- Waga, S. & Stillman, B. (1998) Annu. Rev. Biochem. 67, 721-751. - PubMed
-
- Greider, C. W. & Blackburn, E. H. (1985) Cell 43, 405-413. - PubMed
-
- Singer, M. S. & Gottschling, D. E. (1994) Science 266, 404-409. - PubMed
-
- Cervantes, R. B. & Lundblad, V. (2002) Curr. Opin. Cell Biol. 14, 351-356. - PubMed
-
- McEachern, M. J., Krauskopf, A. & Blackburn, E. H. (2000) Annu. Rev. Genet. 34, 331-358. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
