

Disruption of Ixodes scapularis anticoagulation by using RNA interference
- PMID: 14745044
- PMCID: PMC337020
- DOI: 10.1073/pnas.0307669100
Disruption of Ixodes scapularis anticoagulation by using RNA interference
Abstract
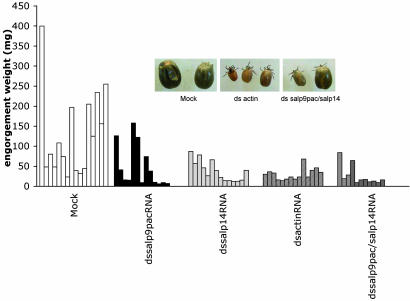
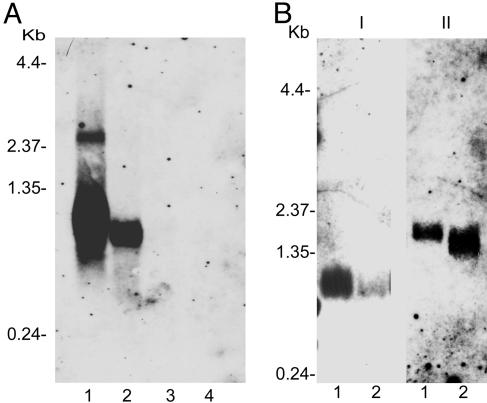
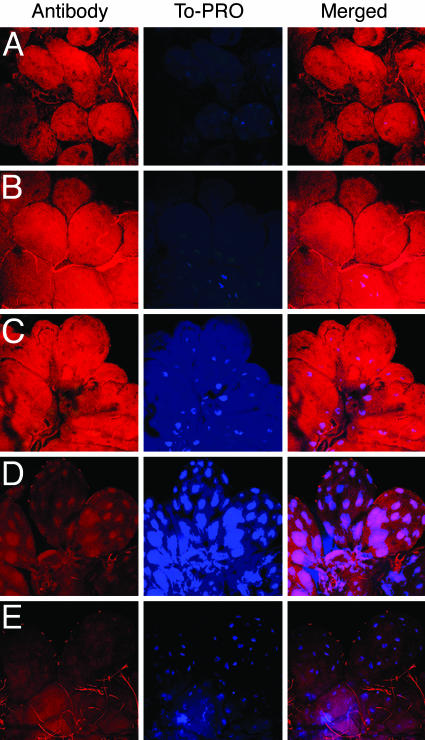
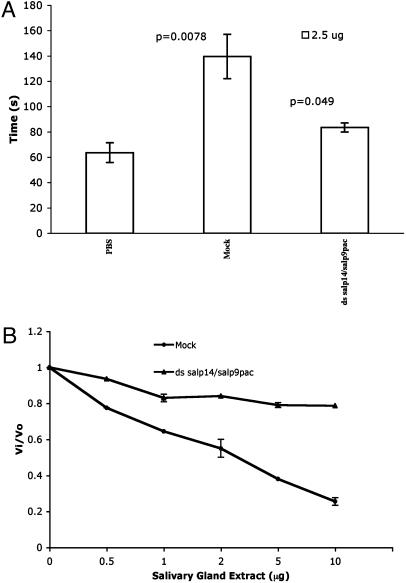
Ixodes scapularis ticks transmit many pathogens, including Borrelia burgdorferi, Anaplasma phagocytophilum, and Babesia microti. Vaccines directed against arthropod proteins injected into the host during tick engorgement could prevent numerous infectious diseases. Salp14, a salivary anticoagulant, poses a key target for such intervention. Salp14 is the prototypic member of a family of potential I. scapularis anticoagulants, expressed and secreted in tick saliva during tick feeding. RNA interference was used to assess the role of Salp14 in tick feeding. Salp14 and its paralogs were silenced, as demonstrated by the reduction of mRNA and protein specific for these antigens. Tick salivary glands lacking Salp14 had reduced anticoagulant activity, as revealed by a 60-80% reduction of anti-factor Xa activity. Silencing the expression of salp14 and its paralogs also reduced the ability of I. scapularis to feed, as demonstrated by a 50-70% decline in the engorgement weights. Because ticks have several anticoagulants, it is likely that the expression of multiple anticoagulants in I. scapularis saliva would have to be ablated simultaneously to abolish tick feeding. These studies demonstrate that RNA interference can silence I. scapularis genes and disrupt their physiologic function in vivo, and they identify vaccine candidates that can alter vector engorgement.
Figures

References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
