

Placental failure and impaired vasculogenesis result in embryonic lethality for neuropathy target esterase-deficient mice
- PMID: 14749382
- PMCID: PMC344166
- DOI: 10.1128/MCB.24.4.1667-1679.2004
Placental failure and impaired vasculogenesis result in embryonic lethality for neuropathy target esterase-deficient mice
Abstract
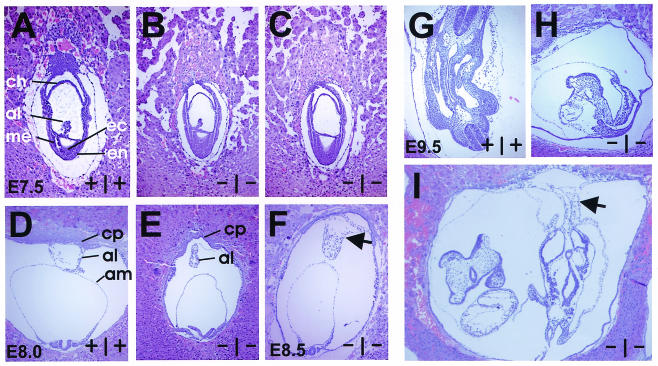
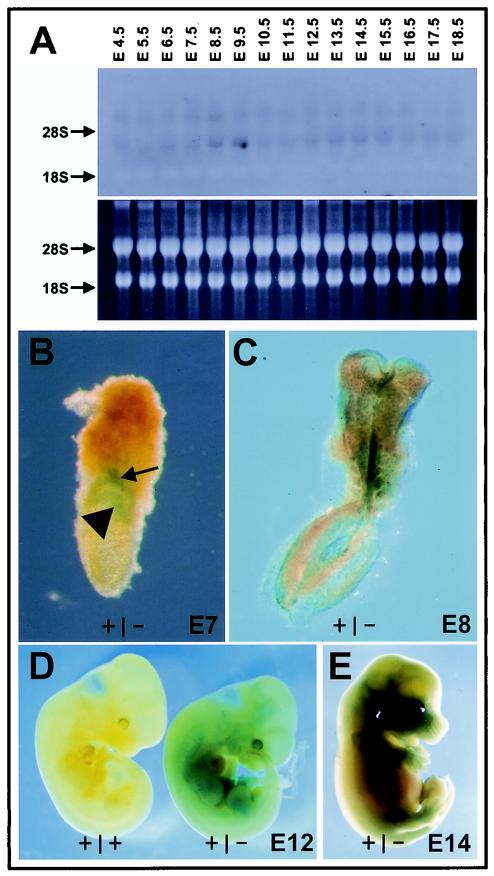
Age-dependent neurodegeneration resulting from widespread apoptosis of neurons and glia characterize the Drosophila Swiss Cheese (SWS) mutant. Neuropathy target esterase (NTE), the vertebrate homologue of SWS, reacts with organophosphates which initiate a syndrome of axonal degeneration. NTE is expressed in neurons and a variety of nonneuronal cell types in adults and fetal mice. To investigate the physiological functions of NTE, we inactivated its gene by targeted mutagenesis in embryonic stem cells. Heterozygous NTE(+/-) mice displayed a 50% reduction in NTE activity but underwent normal organ development. Complete inactivation of the NTE gene resulted in embryonic lethality, which became evident after gastrulation at embryonic day 9 postcoitum (E9). As early as E7.5, mutant embryos revealed growth retardation which did not reflect impaired cell proliferation but rather resulted from failed placental development; as a consequence, massive apoptosis within the developing embryo preceded its resorption. Histological analysis indicated that NTE is essential for the formation of the labyrinth layer and survival and differentiation of secondary giant cells. Additionally, impairment of vasculogenesis in the yolk sacs and embryos of null mutant conceptuses suggested that NTE is also required for normal blood vessel development.
Figures








References
-
- Anson-Cartwright, L., K. Dawson, S. J. Fisher, R. A. Lazzarini, and J. C. Cross. 2000. The glial cells missing-1 protein is essential for branching morphogenesis in the chorioallantoic placenta. Nat. Genet. 25:311-314. - PubMed
-
- Carmeliet, P., V. Ferreira, G. Breier, S. Pollefeyt, L. Kieckens, M. Gertsenstein, M. Fahrig, A. Vandenhoeck, K. Harpal, C. Eberhardt, C. Declercq, J. Pawling, L. Moons, D. Collen, W. Risau, and A. Nagy. 1996. Abnormal blood vessel development and lethality in embryos lacking a single VEGF allele. Nature 380:435-439. - PubMed
-
- Colosi, P., F. Talamantes, and D. I. Linzer. 1987. Molecular cloning and expression of mouse placental lactogen I complementary deoxyribonucleic acid. Mol. Endocrinol. 1:767-776. - PubMed
-
- Cross, J. C., M. L. Flannery, M. A. Blanar, E. Steingrimsson, N. A. Jenkins, N. G. Copeland, W. J. Rutter, and Z. Werb. 1995. Hxt encodes a basic helix-loop-helix transcription factor that regulates trophoblast cell development. Development 121:2513-2523. - PubMed
-
- Ehrich, M., and B. S. Jortner. 2001. Organophosphorus-induced delayed neuropathy, p. 987-1012. In R. Krieger (ed.), Handbook of pesticide toxicology, 2nd ed. Academic Press, San Diego, Calif.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials