

Enhancer I predominance in hepatitis B virus gene expression
- PMID: 14749394
- PMCID: PMC344184
- DOI: 10.1128/MCB.24.4.1799-1808.2004
Enhancer I predominance in hepatitis B virus gene expression
Abstract
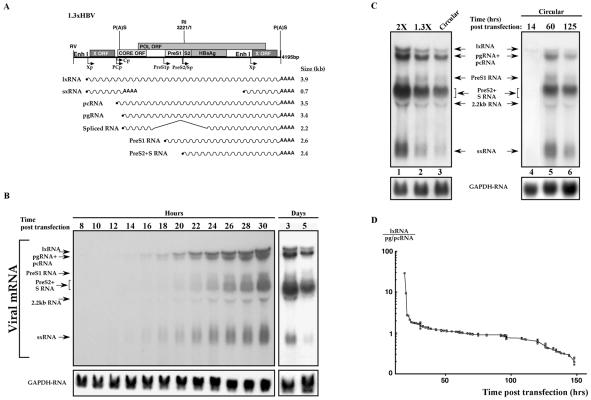
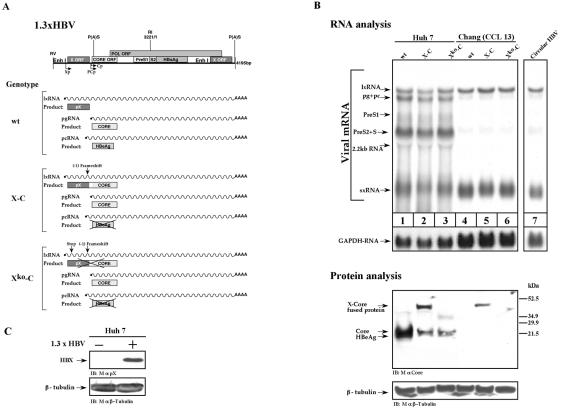
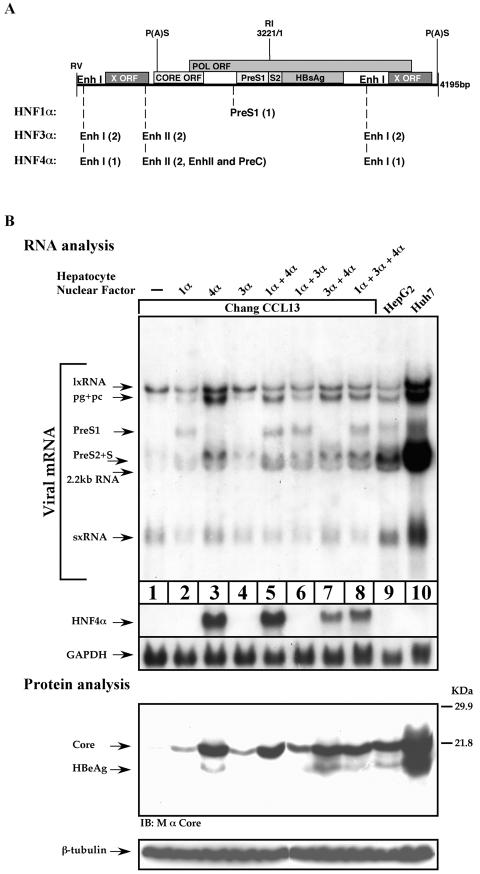
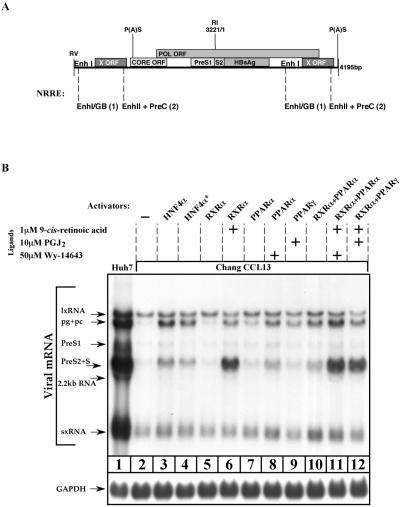
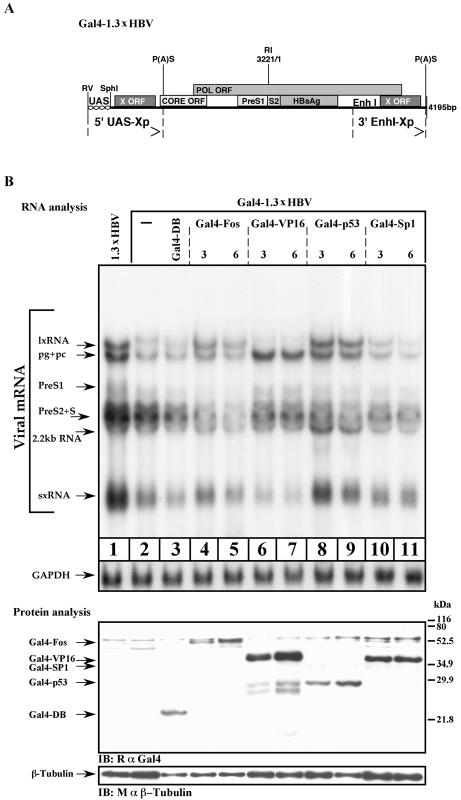
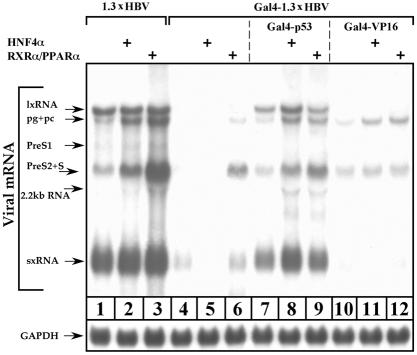
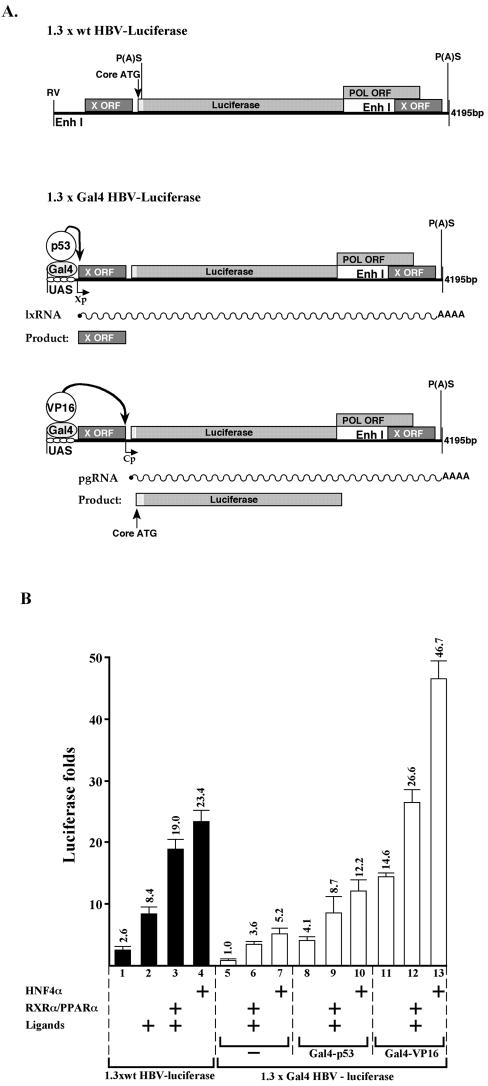
Previous studies of human hepatitis B virus (HBV) transcription revealed the requirement of two enhancer elements. Enhancer I (EnhI) is located upstream of the X promoter and is targeted by multiple activators, including basic leucine zipper proteins, and enhancer II (EnhII) is located upstream to the PreCore promoter and is targeted mainly by nuclear receptors (NRs). The mode of interplay between these enhancers and their unique contributions in regulating HBV transcription remained obscure. By using time course analysis we revealed that the HBV transcripts are categorized into early and late groups. Chang (CCL-13) cells are impaired in expression of the late transcripts. This could be corrected by overexpressing EnhII activators, such as hepatocyte nuclear factor 4 alpha, the retinoid X receptor alpha, and the peroxisome proliferator-activated receptor alpha, suggesting that in Chang cells EnhI but not EnhII is active. Replacing the 5'-end EnhI sequence with a synthetic Gal4 response (UAS) DNA fragment ceased the production of the early transcripts. Under this condition NR overexpression poorly activated EnhII. However, activation of the UAS by Gal4-p53 restored both the expression of the early transcripts and the EnhII response to NRs. Thus, a functional EnhI is required for activation of EnhII. We found a major difference between Gal4-p53 and Gal4-VP16 behavior. Gal4-p53 activated the early transcripts, while Gal4-VP16 inhibited the early transcripts but activated the late transcripts. These findings indicate that the composition of the EnhI binding proteins may play a role in early to late switching. Our data provides strong evidence for the role of EnhI in regulating global and temporal HBV gene expression.
Figures
References
-
- Bock, C. T., S. Kubicka, M. P. Manns, and C. Trautwein. 1999. Two control elements in the hepatitis B virus S-promoter are important for full promoter activity mediated by CCAAT-binding factor. Hepatology 29:1236-1247. - PubMed
-
- Cattaneo, R., H. Will, N. Hernandez, and H. Schaller. 1983. Signals regulating hepatitis B surface antigen transcription. Nature 305:336-338. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous