

Review
doi: 10.1101/gad.1165904.
The Mus81 solution to resolution: generating meiotic crossovers without Holliday junctions
Affiliations
- PMID: 14752007
- PMCID: PMC1851908
- DOI: 10.1101/gad.1165904
Item in Clipboard
Review
The Mus81 solution to resolution: generating meiotic crossovers without Holliday junctions
Genes Dev.
.
No abstract available
Figures
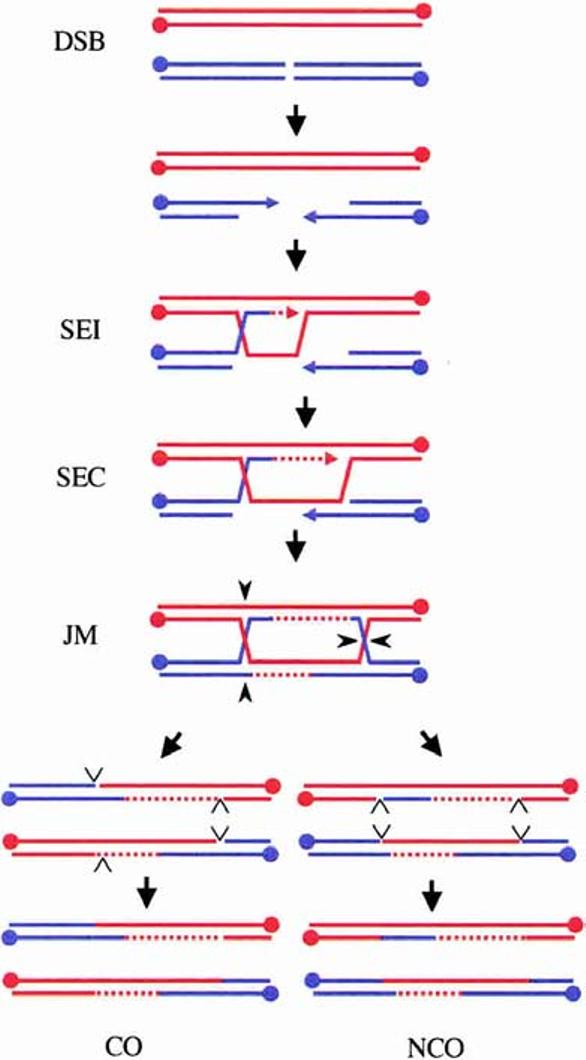
The DSBR model in meiosis. Shown are the duplex DNA strands of two non-sister chromatids from a pair of homologous chromosomes (Homolog 1 is red; Homolog 2 is blue). Meiotic DSBs are repaired following 5′-end resection and the formation of an SEI intermediate. Elongation of the invading strand extends the D-loop until SEC initiates a second round of DNA synthesis. Ligation of the free ends creates a joint molecule containing dHJs. Resolution of the dHJs in an opposite sense (e.g., black carets shown here) leads to the formation of a CO, whereas cleavage in the same sense (e.g., cleavage of the crossed strands) results in an NCO. Resolution consists of two steps: symmetric cleavage to produce duplex DNA with nicks (open carets) and ligation of the nicks. 5′-Ends of the DNA strands are represented by dots, 3′-ends are represented by arrows, and DNA synthesis is indicated by dashed lines.
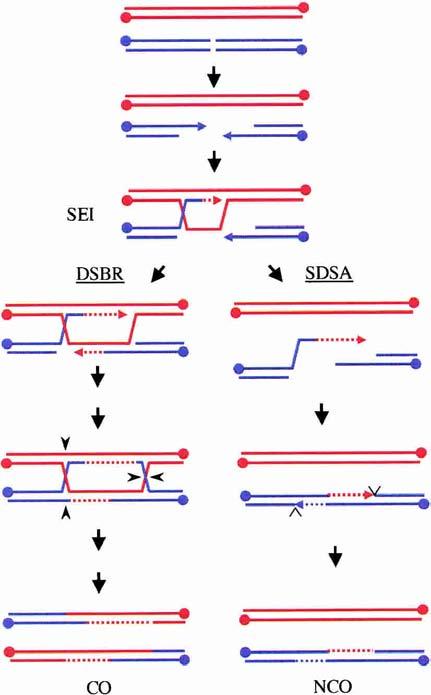
An alternative model for generating NCOs in budding yeast meiosis. Evidence at present suggests that SEI intermediates become committed to forming COs if they are repaired via the DSBR pathway, presumably through a biased resolution of dHJs in the direction of COs (left pathway). The SEI intermediate may be disrupted, however, to allow repair to proceed via the SDSA pathway, resulting in NCO products (right pathway).
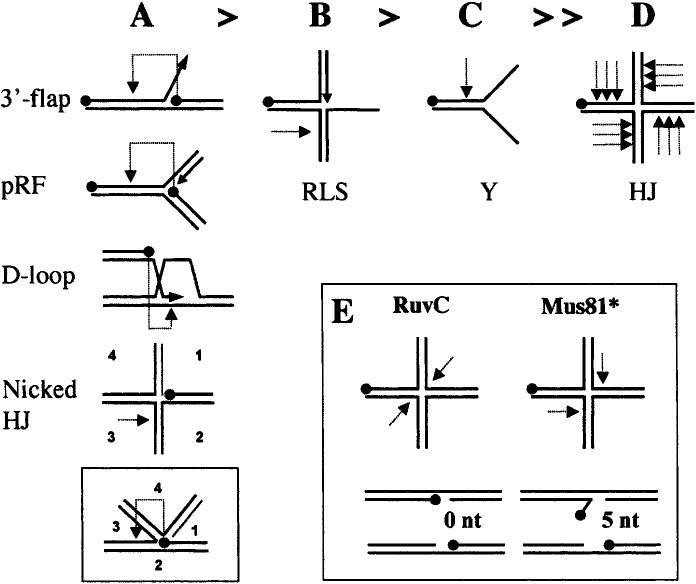
Substrate-specificity of Mus81* in vitro. (A–D) Summary of results from a variety of cleavage studies using rMus81* with substrates presented in order of cleavage efficiency. The most efficient substrates (A) are cleaved well owing to the presence of a 5′-end at their branch junctions (indicated by a black dot). Cleavage of these substrates (light dotted arrow) is directed one-half helical turn upstream of this 5′-end (pRF, pseudoreplication fork). (A, inset) This mechanism is consistent with the cleavage of nicked HJs in the strand opposing the nick. Efficient cleavage of the remaining substrates (B–D) requires an excess of enzyme over substrate. A regressed lagging-strand substrate (RLS, panel B) is cleaved in the opposing strand, while a Y substrate (C) is cleaved upstream of the branch junction. A further increase in enzyme concentration results in nicking of mobile HJs at multiple sites on the 5′-side of the mobile core. (E) Contrast of the HJ cleavage products obtained by symmetrical cleavage with RuvC (nicked duplexes) to the unligatable flap and gap products produced by Mus81*. These 5′-flap products may isomerize into 3′-flaps that are further processed by Mus81* to produce only gapped molecules.
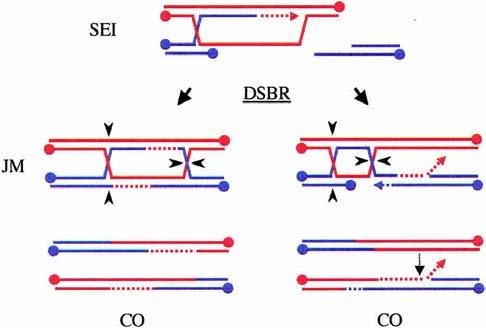
A role for Mus81* in cleaving 3′-flaps. Model II proposes that most meiotic SEI intermediates are processed into COs via the DSBR pathway (left). In a subset of SEI intermediates, however, the invading strand may be elongated excessively (right). In this case, the second strand is captured, not by the D-loop, but by the elongated strand following displacement. Any 3′-flaps that arise from strand annealing are substrates for Mus81* (light dotted arrow). In this pathway, heteroduplex DNA is found adjacent to the dHJs rather than between them.
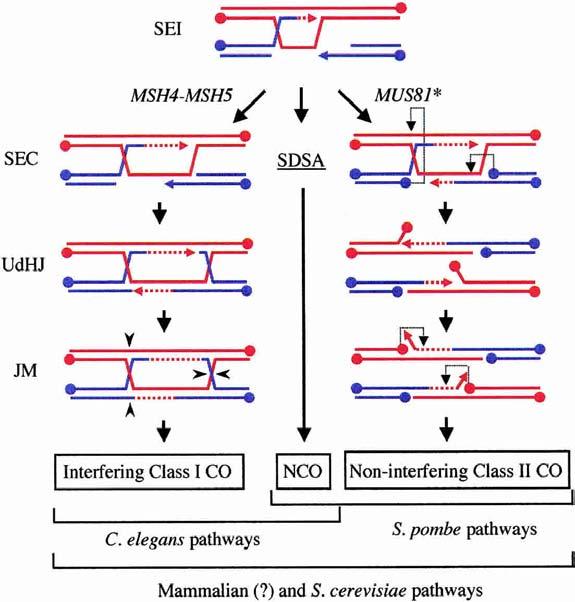
The role of Mus81* in creating COs without HJs. Model III proposes that meiotic SEI intermediates have three fates depending on the organism and pathway choice. All organisms use SDSA to produce NCOs (center). Class I interfering COs occur in organisms containing MSH4–MSH5 and arise from the DSBR pathway (left). Class II noninterfering COs are proposed to arise from the action of Mus81* on an SEC intermediate (right). The SEC intermediate provides targets for Mus81* cleavage at the D-loop and at the half-junction (light dotted arrows). Cleavage at these sites results in an obligatory CO product containing 5′-flaps and gaps. Here the flaps are shown to isomerize into 3′-flaps prior to cleavage by a second round of Mus81* action. Filling-in of the gaps and ligation completes the CO product (not shown).
References
-
- Agarwal S, Roeder GS. Zip3 provides a link between recombination enzymes and synaptonemal complex proteins. Cell. 2000;102:245–255. - PubMed
-
- Alani E, Lee S, Kane MF, Griffith J, Kolodner RD. Saccharomyces cerevisiae MSH2, a mispaired base recognition protein, also recognizes Holliday junctions in DNA. J. Mol. Biol. 1997;265:289–301. - PubMed
-
- Allers T, Lichten M. Differential timing and control of noncrossover and crossover recombination during meiosis. Cell. 2001a;106:47–57. - PubMed
-
- Allers T, Lichten M. Intermediates of yeast meiotic recombination contain heteroduplex DNA. Mol. Cell. 2001b;8:225–231. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases