

doi: 10.1101/gad.282604.
Roles of Bifocal, Homer, and F-actin in anchoring Oskar to the posterior cortex of Drosophila oocytes
Affiliations
- PMID: 14752008
- PMCID: PMC324420
- DOI: 10.1101/gad.282604
Item in Clipboard
Roles of Bifocal, Homer, and F-actin in anchoring Oskar to the posterior cortex of Drosophila oocytes
Genes Dev.
.
Abstract
Transport, translation, and anchoring of osk mRNA and proteins are essential for posterior patterning of Drosophila embryos. Here we show that Homer and Bifocal act redundantly to promote posterior anchoring of the osk gene products. Disruption of actin microfilaments, which causes delocalization of Bifocal but not Homer from the oocyte cortex, severely disrupts anchoring of osk gene products only when Homer (not Bifocal) is absent. Our data suggest that two processes, one requiring Bifocal and an intact F-actin cytoskeleton and a second requiring Homer but independent of intact F-actin, may act redundantly to mediate posterior anchoring of the osk gene products.
Figures
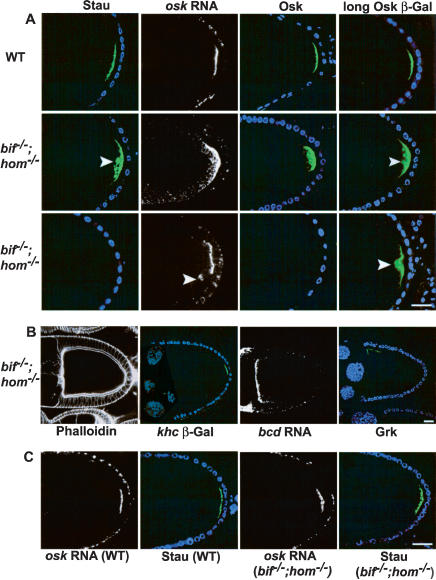
Loss of both bifocal and homer causes defective anchoring of the posterior determinants in oocytes. (A) Stage 10 wild-type and bif;hom double mutant oocytes showing the localization of Stau (green), osk RNA (white), Osk (green), and Osk-βGal (green). Two sets of double mutant oocytes are shown (second and third rows) to illustrate the different severity of the phenotypes observed; for example, Stau and Osk can either be loosely cytoplasmic (second row) or undetectable (third row). Arrowheads point to sites of cytoplasmic localization. DNA staining (blue) marks the position of follicle cell nuclei. (B) Double mutant oocytes assessed with phalloidin, khc β-Gal, bcd RNA in situ, and anti-Grk, all of which appear normal. (C) osk RNA and Stau localization in wild-type and double mutant stage 9 oocytes; the normal localization seen at stage 9 indicates that the defects seen in stage 10 double mutant oocytes are due to defects in anchoring. (Bar length is 10 μm in all figures.)
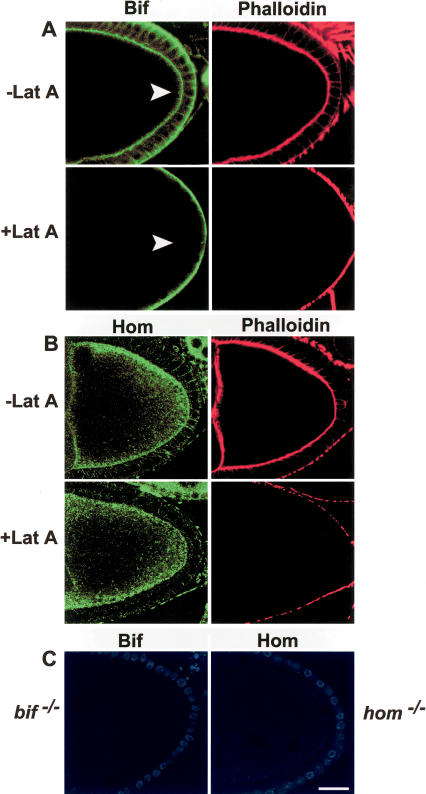
Bifocal and Homer localization in the presence and absence of intact microfilaments. (A) Anti-Bif (green) and phalloidin (red) staining of Lat A-treated and untreated wild-type oocytes. Note that cortical Bif staining (arrowheads) in the oocyte disappears in the absence of an intact F-actin cytoskeleton. (B) Anti-Hom (green) and phalloidin (red) staining in the presence and absence of Lat A treatment. Note that Hom staining at the cortex of the oocyte remains even in the absence of an intact F-actin cytoskeleton. (C) Anti-Bif and anti-Hom staining of their respective mutant oocytes. In these oocytes the follicle cell nuclei are in blue. Note that in Lat A-treated ovaries, residual phalloidin staining remains on the exterior border of the follicle cells, whereas it is totally absent from the oocyte cortex, perhaps due to a lower actin turnover in the follicle cells.
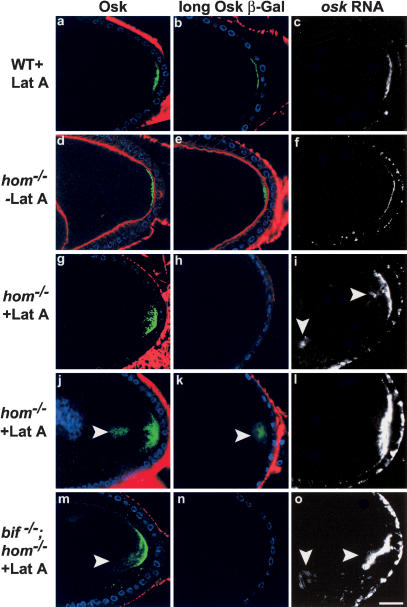
Osk protein and RNA localization in wild-type and hom oocytes in the presence or absence of intact microfilaments. In the great majority of wild-type stage-10 oocytes (a-c), in which the cortical F-actin (phalloidin, red) has been rendered undetectable by Lat A treatment, Osk (green, a), Osk-βGal (green, b), and osk RNA (white, c) are localized as a tight posterior crescent similar to that of untreated wild-type oocytes (see Table 1 for quantitation). DNA staining shows the position of the follicle cell nuclei (blue). In hom oocytes not treated with Lat A (d-f), cortical F-actin (red) can be seen, and Osk protein (d), Osk-βGal (e), and osk RNA (f) localizations are wild type. In Lat A-treated hom oocytes (g-l), the cortical F-actin is undetectable. osk RNA and proteins show diffuse (g,i,j,l) and in some cases prominent cytoplasmic localization (arrowheads, i-k). In several cases the Osk-βGal line does not show any staining in hom mutants treated with Lat A (example in h). Similar results are seen with Lat A treatment of hom;bif double mutants (m-o; cytoplasmic localization of Osk and osk RNA are indicated by arrowheads in m,o).
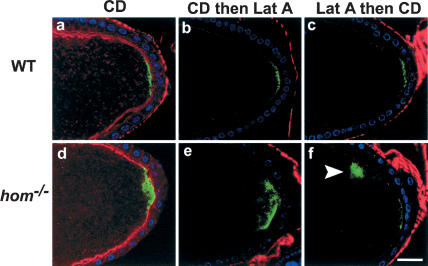
Osk localization in drug-treated wild-type and hom mutants and wild-type stage-9 and -10 oocytes treated with CD alone (a: n = 23, 26% show diffuse staining, whereas the others are normal), CD followed by Lat A (b: n = 18, 22% show diffuse staining), or Lat A followed by CD (c: n = 15, 20% show diffuse staining). Phalloidin staining in these oocytes is in red. Osk protein (green) in these oocytes was detected using anti-Osk antibody. The protein localization in most cases is still predominantly at the posterior cortex, as in the wild-type oocytes. Similar experiments with hom mutant oocytes treated with CD alone (d: 27.2% show diffuse staining; 45.5%, cytoplasmic staining, n = 11), CD followed by Lat A (e: 22.2%, diffuse staining; 16.7%, no staining; 44.4%, cytoplasmic staining, n = 18), and Lat A followed by CD (f: 50%, diffuse staining; 10%, no stain; 30%, cytoplasmic staining, n = 10) all show enhanced diffuse or cytoplasmic staining (arrowhead in f) of Osk after drug treatment. DNA staining (blue) shows the position of the follicle cell nuclei.
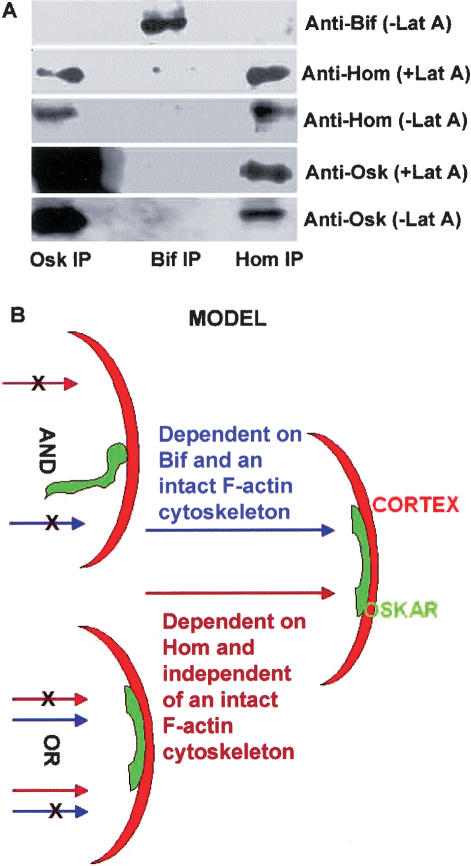
Immunoprecipitations and proposed model. (A) Western blot analysis of co-immunoprecipitations (co-IPs) from extracts of ovaries treated or not treated with Lat A. Anti-Osk, anti-Bif, and anti-Hom co-IPs were probed with the antibodies indicated. Osk appears to complex with Hom in extracts derived from Lat A-treated and untreated ovaries. Bif does not co-IP with Osk or Hom and serves as a control. Note that the amount of Osk brought down in the Osk IP with Lat A is similar to the amount of Osk brought down in the absence of Lat A. (B) A schematic model proposing that the posterior cortical anchoring of the osk gene products in the oocyte is mediated by an F-actin-dependent pathway requiring bif and a second parallel pathway requiring hom which does not require intact F-actin; either pathway can promote anchoring of the osk gene products at the posterior cortex of the great majority of oocytes. Posterior is toward the right.
References
-
- Brakeman P.R., Lanahan, A.A., O'Brien, R., Roche, K., Barnes, C.A., Huganir, R.L., and Worley, P.F. 1997. Homer: A protein that selectively binds metabotropic glutamate receptors. Nature 386: 284-288. - PubMed
-
- Broadus J. and Doe, C.Q. 1997. Extrinsic cues, intrinsic cues and microfilaments regulate asymmetric protein localization in Drosophila neuroblasts. Curr. Biol. 7: 827-835. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases