

Splicing enhances translation in mammalian cells: an additional function of the exon junction complex
- PMID: 14752011
- PMCID: PMC324426
- DOI: 10.1101/gad.1163204
Splicing enhances translation in mammalian cells: an additional function of the exon junction complex
Abstract
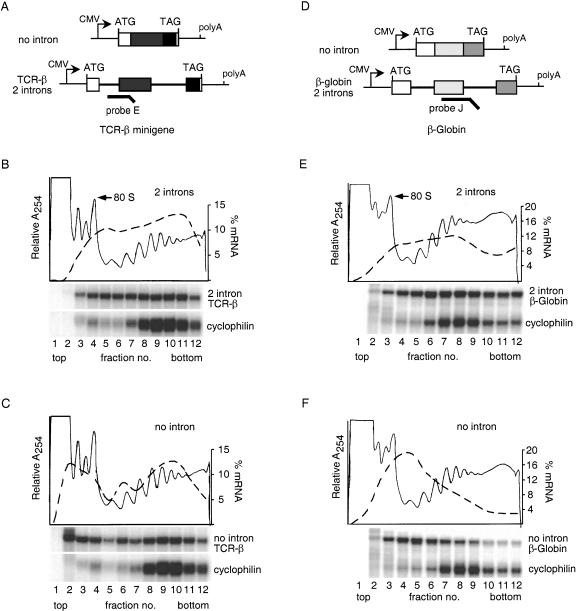
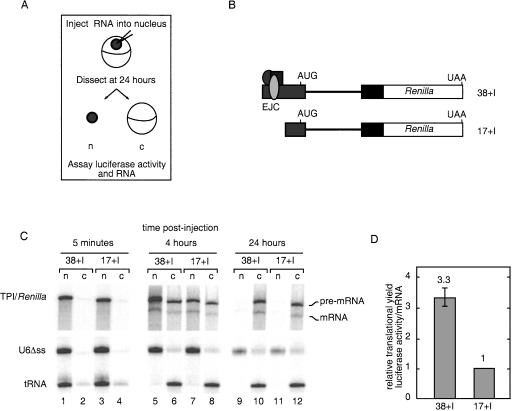
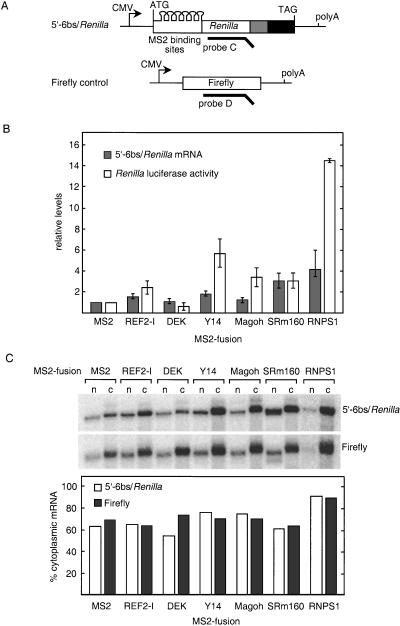
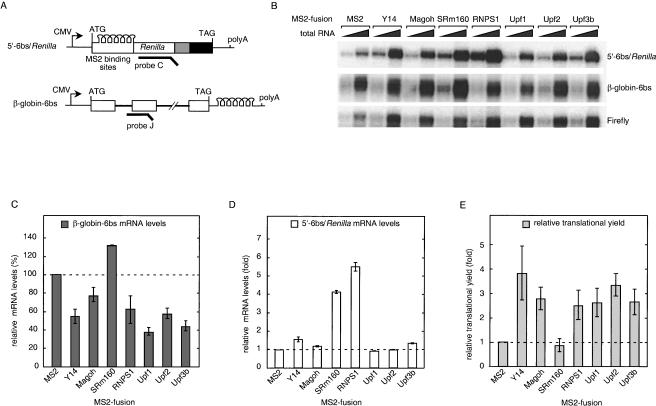
In mammalian cells, spliced mRNAs yield greater quantities of protein per mRNA molecule than do otherwise identical mRNAs not made by splicing. This increased translational yield correlates with enhanced cytoplasmic polysome association of spliced mRNAs, and is attributable to deposition of exon junction complexes (EJCs). Translational stimulation can be replicated by tethering the EJC proteins Y14, Magoh, and RNPS1 or the nonsense-mediated decay (NMD) factors Upf1, Upf2, and Upf3b to an intronless reporter mRNA. Thus, in addition to its previously characterized role in NMD, the EJC also promotes mRNA polysome association. Furthermore, the ability to stimulate translation when bound inside an open reading frame appears to be a general feature of factors required for NMD.
Figures

References
-
- Badolato J., Gardiner, E., Morrison, N., and Eisman, J. 1995. Identification and characterization of a novel human RNA-binding protein. Gene 166: 323-327. - PubMed
-
- Bouvet P. and Wolffe, A.P. 1994. A role for transcription and FRGY2 in masking maternal mRNA within Xenopus oocytes. Cell 77: 931-941. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases