

A novel LIM protein Cal promotes cardiac differentiation by association with CSX/NKX2-5
- PMID: 14757752
- PMCID: PMC2172236
- DOI: 10.1083/jcb.200309159
A novel LIM protein Cal promotes cardiac differentiation by association with CSX/NKX2-5
Abstract
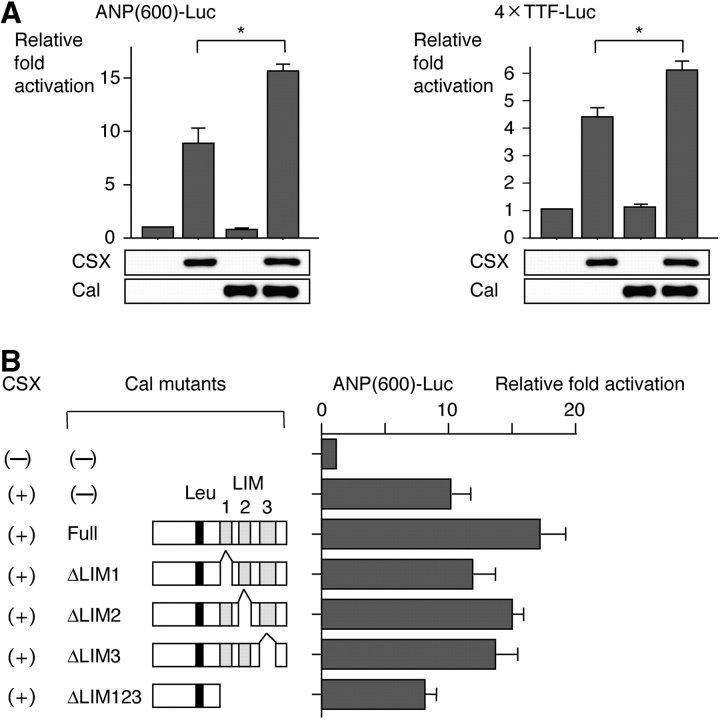
The cardiac homeobox transcription factor CSX/NKX2-5 plays an important role in vertebrate heart development. Using a yeast two-hybrid screening, we identified a novel LIM domain-containing protein, named CSX-associated LIM protein (Cal), that interacts with CSX/NKX2-5. CSX/NKX2-5 and Cal associate with each other both in vivo and in vitro, and the LIM domains of Cal and the homeodomain of CSX/NKX2-5 were necessary for mutual binding. Cal itself possessed the transcription-promoting activity, and cotransfection of Cal enhanced CSX/NKX2-5-induced activation of atrial natriuretic peptide gene promoter. Cal contained a functional nuclear export signal and shuttled from the cytoplasm into the nucleus in response to calcium. Accumulation of Cal in the nucleus of P19CL6 cells promoted myocardial cell differentiation accompanied by increased expression levels of the target genes of CSX/NKX2-5. These results suggest that a novel LIM protein Cal induces cardiomyocyte differentiation through its dynamic intracellular shuttling and association with CSX/NKX2-5.
Copyright The Rockefeller University Press
Figures











Similar articles
-
Characterization of homo- and heterodimerization of cardiac Csx/Nkx2.5 homeoprotein.J Biol Chem. 2001 Feb 16;276(7):4570-80. doi: 10.1074/jbc.M004995200. Epub 2000 Oct 20. J Biol Chem. 2001. PMID: 11042197
-
Up-regulation of natriuretic peptides in the ventricle of Csx/Nkx2-5 transgenic mice.Biochem Biophys Res Commun. 2000 Apr 21;270(3):1074-9. doi: 10.1006/bbrc.2000.2561. Biochem Biophys Res Commun. 2000. PMID: 10772952
-
Dual effects of the homeobox transcription factor Csx/Nkx2-5 on cardiomyocytes.Biochem Biophys Res Commun. 2002 Nov 8;298(4):493-500. doi: 10.1016/s0006-291x(02)02497-x. Biochem Biophys Res Commun. 2002. PMID: 12408979
-
Cardiac transcription factor Csx/Nkx2-5: Its role in cardiac development and diseases.Pharmacol Ther. 2005 Aug;107(2):252-68. doi: 10.1016/j.pharmthera.2005.03.005. Pharmacol Ther. 2005. PMID: 15925411 Review.
-
CAMTA in cardiac hypertrophy.Cell. 2006 May 5;125(3):427-9. doi: 10.1016/j.cell.2006.04.015. Cell. 2006. PMID: 16678087 Review.
Cited by
-
Migfilin: Cell Adhesion Effect and Comorbidities.Onco Targets Ther. 2022 Apr 19;15:411-422. doi: 10.2147/OTT.S357355. eCollection 2022. Onco Targets Ther. 2022. PMID: 35469339 Free PMC article. Review.
-
Migfilin, a molecular switch in regulation of integrin activation.J Biol Chem. 2009 Feb 13;284(7):4713-22. doi: 10.1074/jbc.M807719200. Epub 2008 Dec 13. J Biol Chem. 2009. PMID: 19074766 Free PMC article.
-
Migfilin and filamin as regulators of integrin activation in endothelial cells and neutrophils.PLoS One. 2011;6(10):e26355. doi: 10.1371/journal.pone.0026355. Epub 2011 Oct 17. PLoS One. 2011. PMID: 22043318 Free PMC article.
-
Migfilin promotes autophagic flux through direct interaction with SNAP29 and Vamp8.J Cell Biol. 2024 Nov 4;223(11):e202312119. doi: 10.1083/jcb.202312119. Epub 2024 Sep 16. J Cell Biol. 2024. PMID: 39283311 Free PMC article.
-
Migfilin protein promotes migration and invasion in human glioma through epidermal growth factor receptor-mediated phospholipase C-γ and STAT3 protein signaling pathways.J Biol Chem. 2012 Sep 21;287(39):32394-405. doi: 10.1074/jbc.M112.393900. Epub 2012 Jul 25. J Biol Chem. 2012. PMID: 22843679 Free PMC article.
References
-
- Akazawa, H., and I. Komuro. 2003. Roles of cardiac transcription factors in cardiac hypertrophy. Circ. Res. 92:1079–1088. - PubMed
-
- Akazawa, H., I. Komuro, Y. Sugitani, Y. Yazaki, R. Nagai, and T. Noda. 2000. Targeted disruption of the homeobox transcription factor Bapx1 results in lethal skeletal dysplasia with asplenia and gastroduodenal malformation. Genes Cells. 5:499–513. - PubMed
-
- Arber, S., and P. Caroni. 1996. Specificity of single LIM motifs in targeting and LIM/LIM interactions in situ. Genes Dev. 10:289–300. - PubMed
-
- Arber, S., G. Halder, and P. Caroni. 1994. Muscle LIM protein, a novel essential regulator of myogenesis, promotes myogenic differentiation. Cell. 79:221–231. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
