

Intracranial self-administration of ethanol within the ventral tegmental area of male Wistar rats: evidence for involvement of dopamine neurons
- PMID: 14762123
- PMCID: PMC6793578
- DOI: 10.1523/JNEUROSCI.1319-03.2004
Intracranial self-administration of ethanol within the ventral tegmental area of male Wistar rats: evidence for involvement of dopamine neurons
Abstract
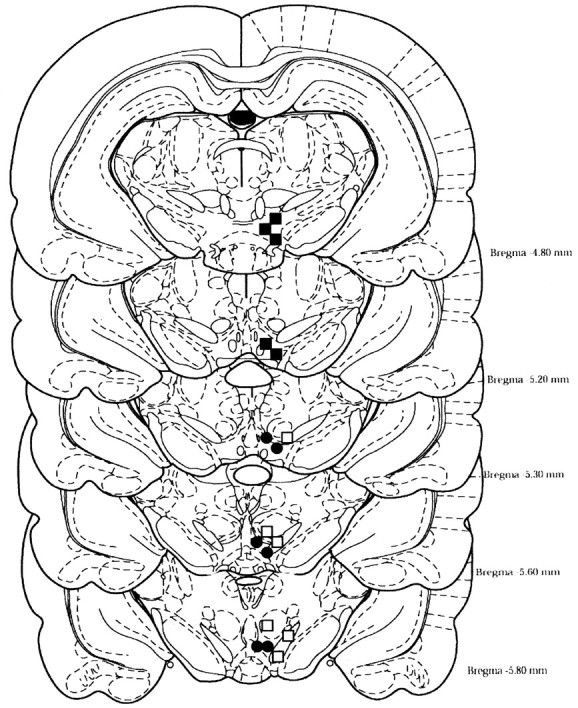
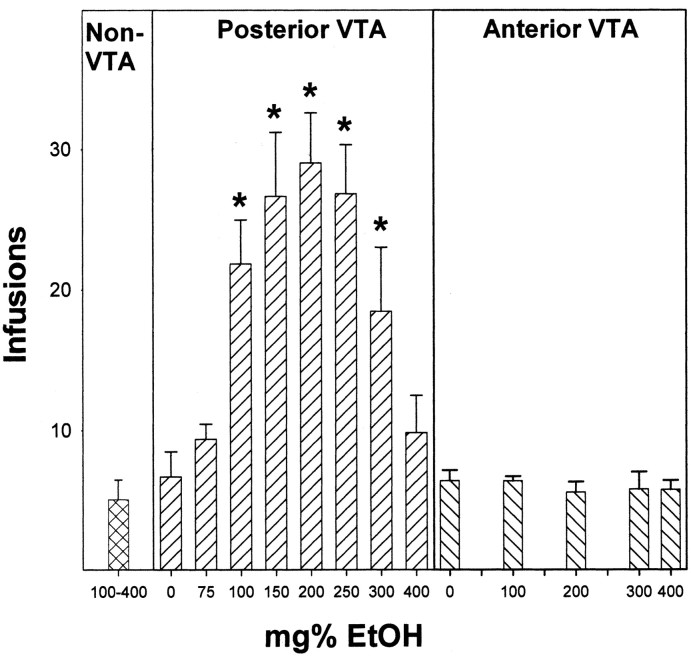
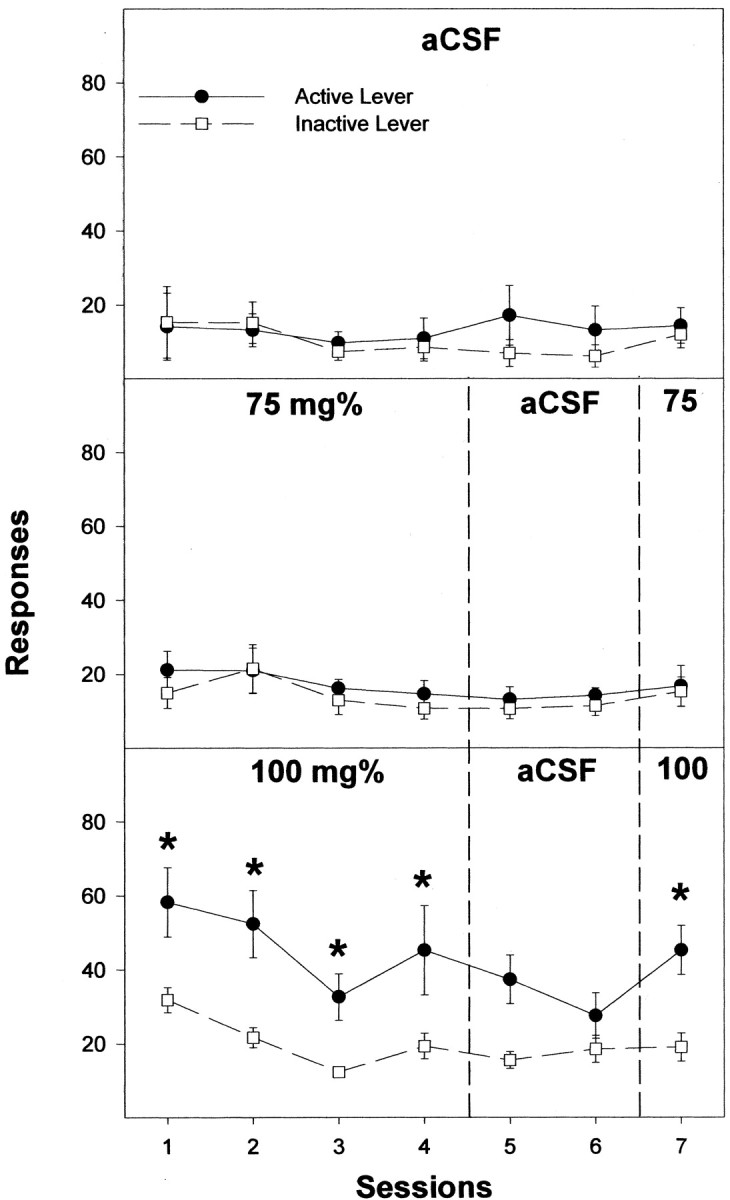
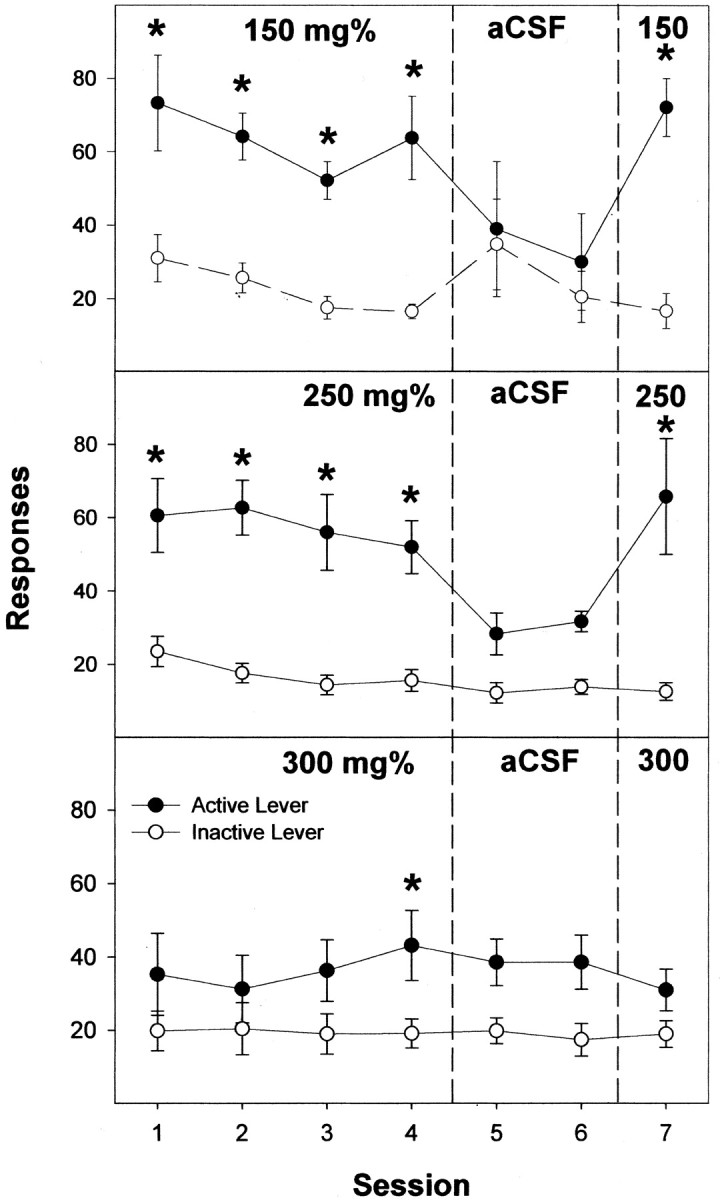
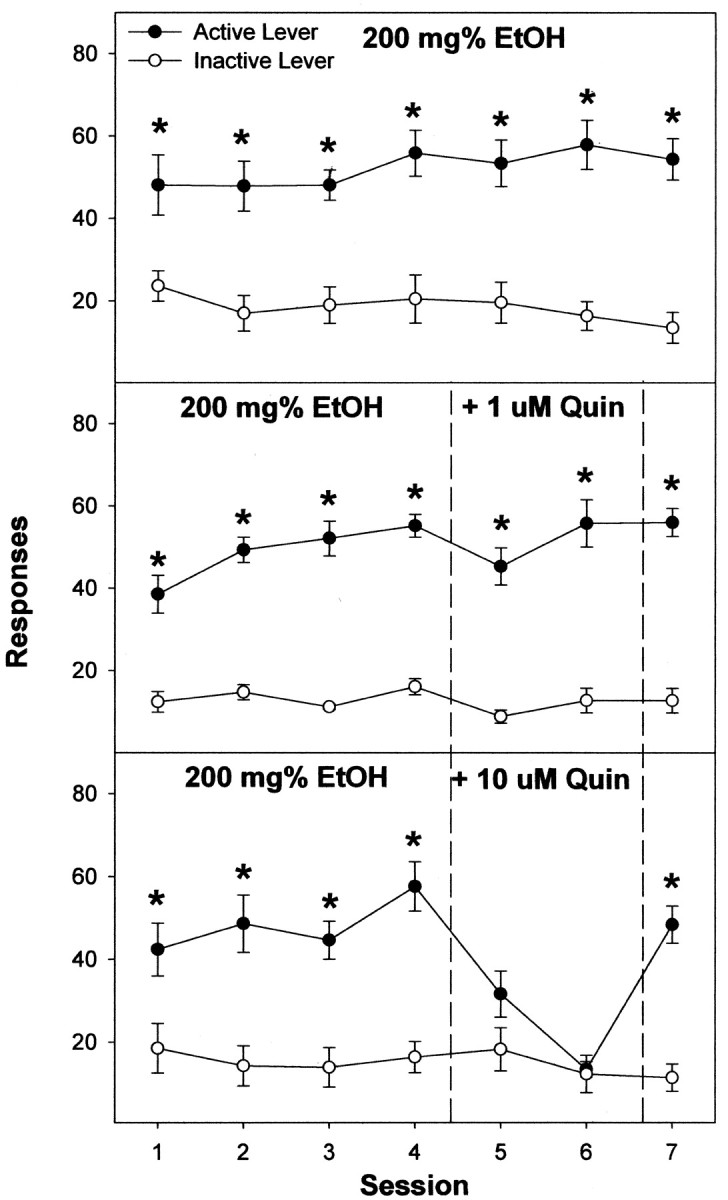
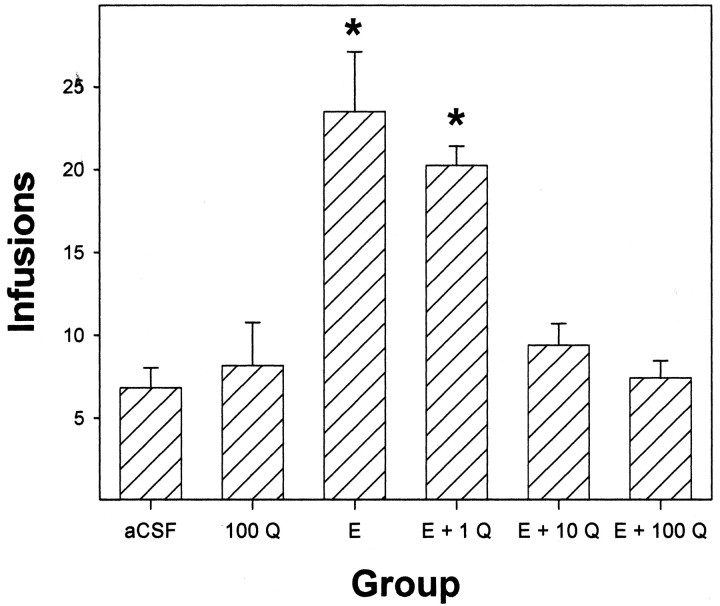
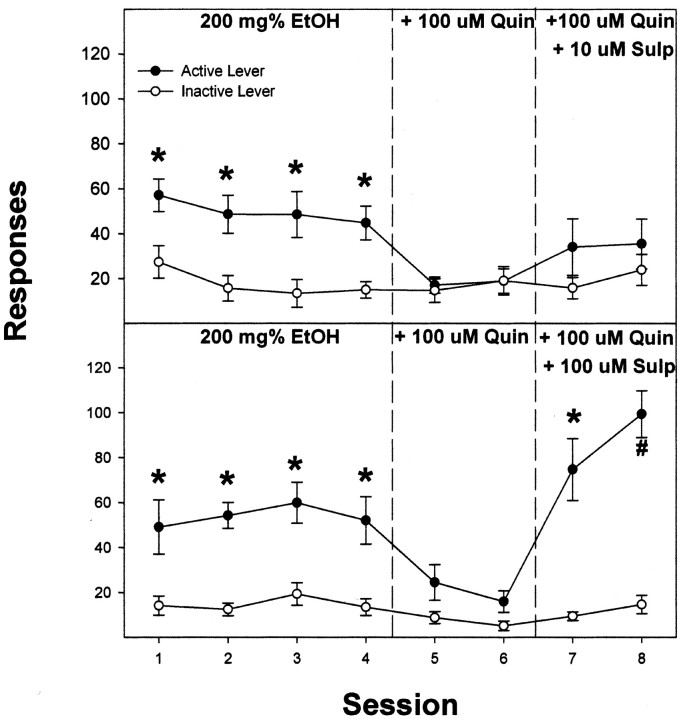
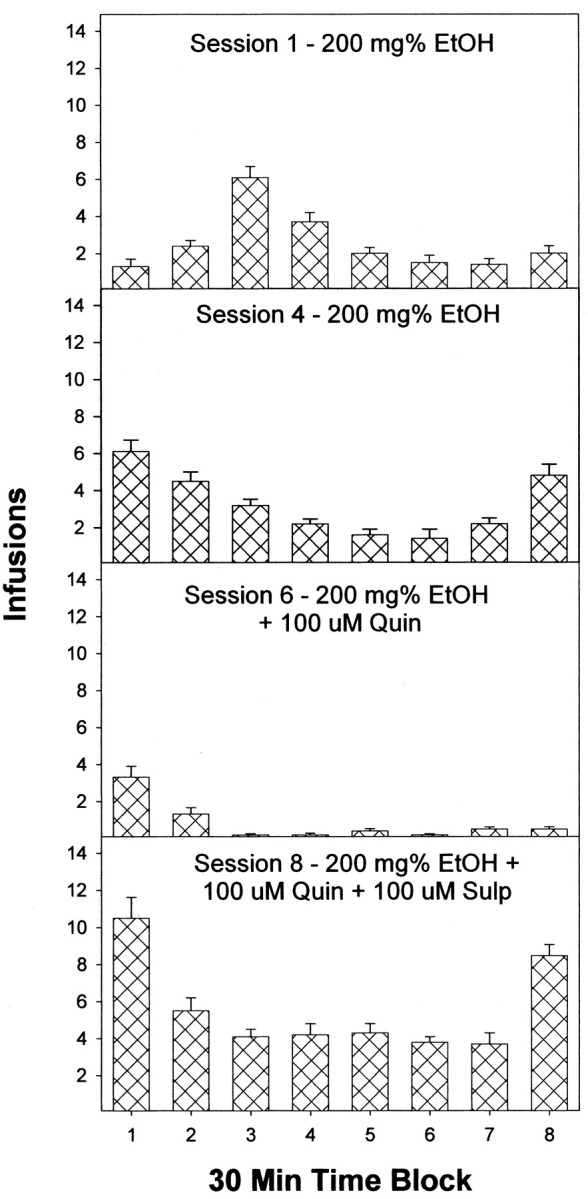
Previous work from our laboratory indicated that female Wistar rats will self-administer ethanol (EtOH) directly into the posterior ventral tegmental area (VTA). These results suggested that VTA dopamine (DA) neurons might be involved in mediating the reinforcing actions of EtOH within this region. The objectives of this study were to determine (1) the dose-response effects for the self-administration of EtOH into the VTA of male Wistar rats, and (2) the involvement of VTA DA neurons in the reinforcing actions of EtOH within the VTA. Adult male Wistar rats were implanted stereotaxically with guide cannulas aimed at the posterior or anterior VTA. After 1 week, rats were placed in standard two-lever (active and inactive) experimental chambers for a total of seven to eight sessions. The first experiment determined the intracranial self-administration of EtOH (0-400 mg%) into the posterior and anterior VTA. The second experiment examined the effects of coadministration of the D2/3 agonist quinpirole on the acquisition and maintenance of EtOH self-infusions into the posterior VTA. The final experiment determined the effects of a D2 antagonist (sulpiride) to reinstate self-administration behavior in rats given EtOH and quinpirole to coadminister. Male Wistar rats self-infused 100-300 mg% EtOH directly into the posterior, but not anterior, VTA. Coadministration of quinpirole prevented the acquisition and extinguished the maintenance of EtOH self-infusion into the posterior VTA, and addition of sulpiride reinstated EtOH self-administration. The results of this study indicate that EtOH is reinforcing within the posterior VTA of male Wistar rats and suggest that activation of VTA DA neurons is involved in this process.
Figures
References
-
- Arnt J, Scheel-Kruger J (1979) GABA in the ventral tegmental area: differential regional effects on locomotion, aggression and food intake after microinjection of GABA agonists and antagonists. Life Sci 25: 1351–1360. - PubMed
-
- Becker JB (1999) Gender differences in dopaminergic function in striatum and nucleus accumbens. Pharmacol Biochem Behav 64: 803–812. - PubMed
-
- Boehm II SL, Piercy MM, Bergstrom HC, Phillips TJ (2002) Ventral tegmental area region governs GABA(B) receptor modulation of ethanol-stimulated activity in mice. Neuroscience 115: 185–200. - PubMed
-
- Bozarth MA, Wise RA (1980) Electrolytic microinfusion transducer system: an alternative method of intra-cranial drug application. J Neurosci Methods 2: 273–275. - PubMed
-
- Bozarth MA, Wise RA (1981) Intracranial self-administration of morphine into the ventral tegmental area in rats. Life Sci 28: 551–555. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources