

Crystal structure of a myristoylated CAP-23/NAP-22 N-terminal domain complexed with Ca2+/calmodulin
- PMID: 14765114
- PMCID: PMC381001
- DOI: 10.1038/sj.emboj.7600093
Crystal structure of a myristoylated CAP-23/NAP-22 N-terminal domain complexed with Ca2+/calmodulin
Abstract
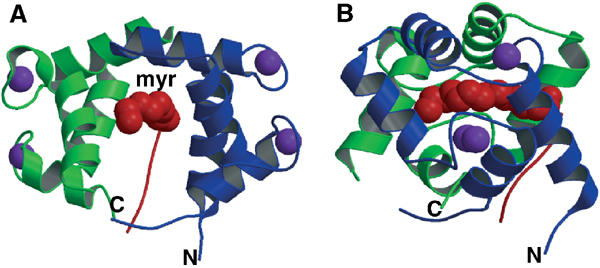
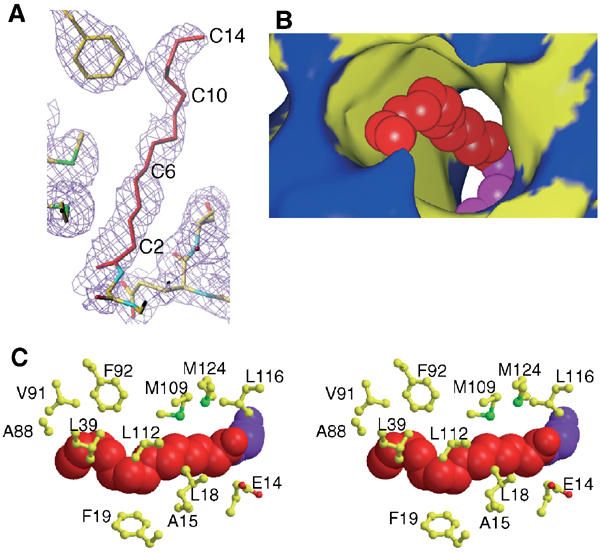
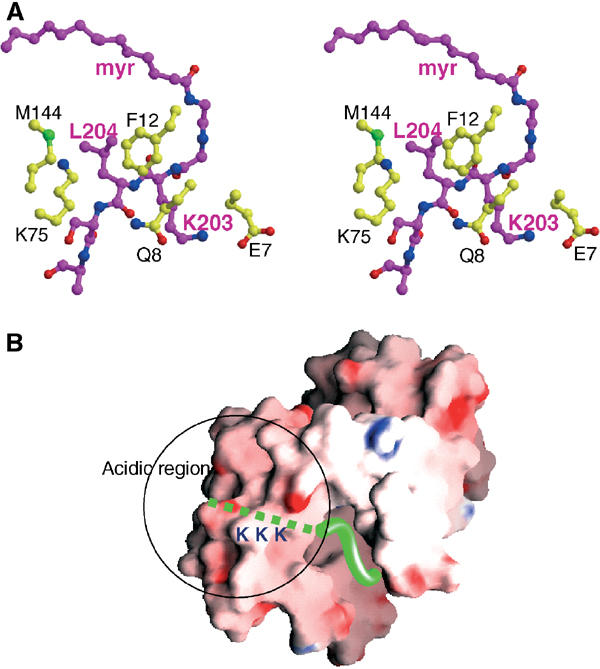
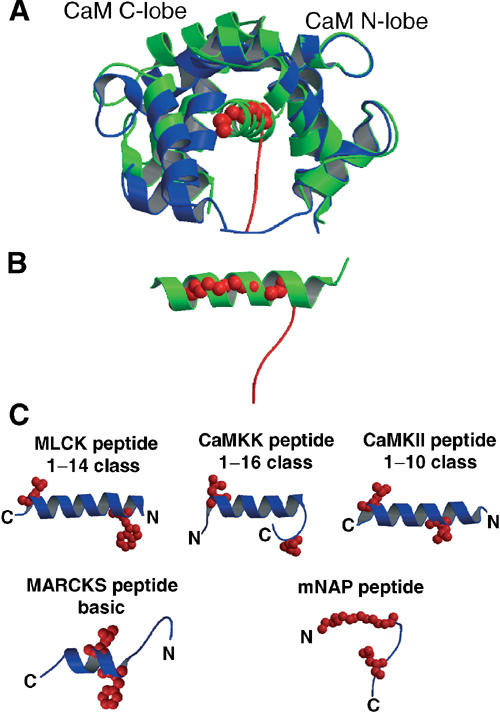
A variety of viral and signal transduction proteins are known to be myristoylated. Although the role of myristoylation in protein-lipid interaction is well established, the involvement of myristoylation in protein-protein interactions is less well understood. CAP-23/NAP-22 is a brain-specific protein kinase C substrate protein that is involved in axon regeneration. Although the protein lacks any canonical calmodulin (CaM)-binding domain, it binds CaM with high affinity. The binding of CAP-23/NAP-22 to CaM is myristoylation dependent and the N-terminal myristoyl group is directly involved in the protein-protein interaction. Here we show the crystal structure of Ca2+-CaM bound to a myristoylated peptide corresponding to the N-terminal domain of CAP-23/NAP-22. The myristoyl moiety of the peptide goes through a hydrophobic tunnel created by the hydrophobic pockets in the N- and C-terminal domains of CaM. In addition to the myristoyl group, several amino-acid residues in the peptide are important for CaM binding. This is a novel mode of binding and is very different from the mechanism of binding in other CaM-target complexes.
Figures
References
-
- Ames JB, Ishima R, Tanaka T, Gordon JI, Stryer L, Ikura M (1997) Molecular mechanics of calcium-myristoyl switches. Nature 389: 198–202 - PubMed
-
- Bomze HM, Bulsara KR, Iskandar BJ, Caroni P, Skene JHP (2001) Spinal axon regeneration evoked by replacing two growth cone proteins in adult neurons. Nat Neurosci 4: 38–43 - PubMed
-
- Brunger A, Adams PD, Clore GM, DeLano WL, Gros P, Grosse-Kunstleve RW, Jiang JS, Kuszewski J, Nilges M, Pannu NS, Read RJ, Rice LM, Simonson T, Warren GL (1998) Crystallography and NMR system: a new software suite for macromolecular structure determination. Acta Crystallogr D 54: 905–921 - PubMed
-
- CCP4 (1994) The CCP4 suite: programs for protein crystallography. Acta Crystallogr D 50: 760–763 - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
