

An extracellular aspartic protease functions in Arabidopsis disease resistance signaling
- PMID: 14765119
- PMCID: PMC380998
- DOI: 10.1038/sj.emboj.7600086
An extracellular aspartic protease functions in Arabidopsis disease resistance signaling
Abstract
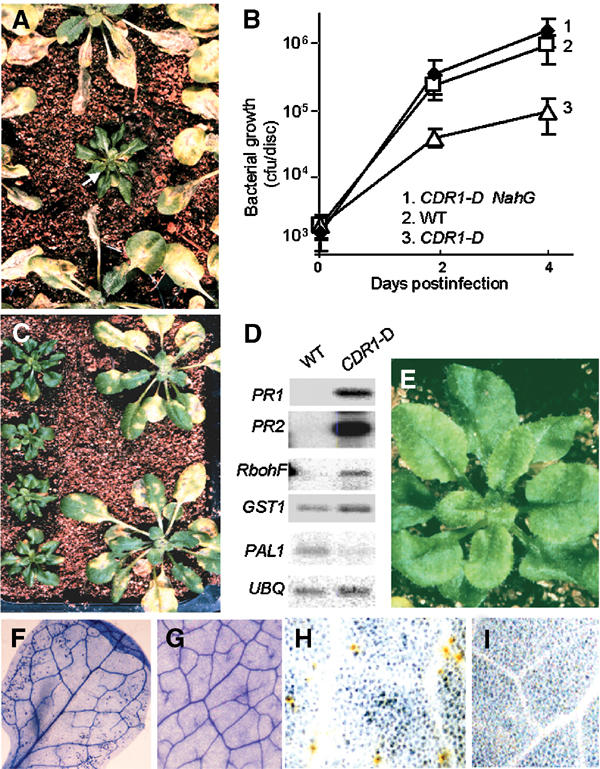
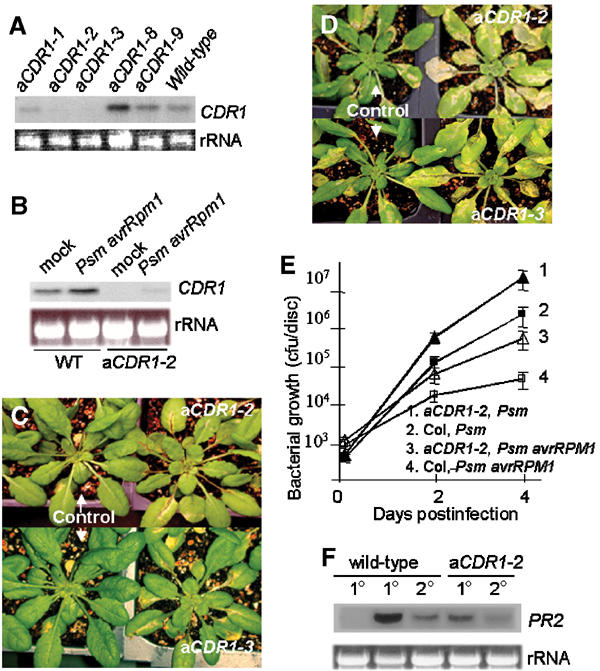
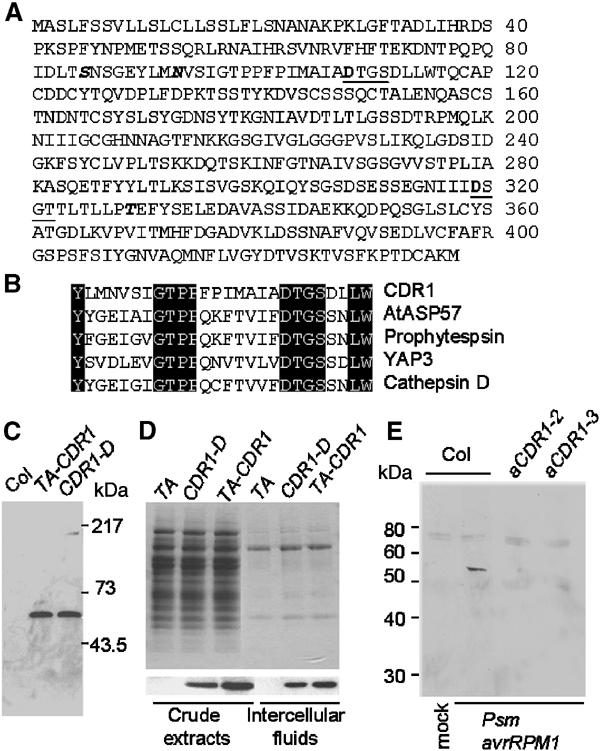
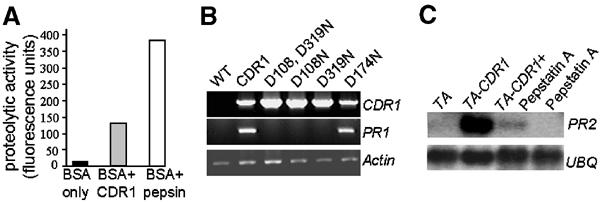
We have used activation tagging with T-DNA carrying cauliflower mosaic virus 35S enhancers to investigate the complex signaling networks underlying disease resistance in Arabidopsis. From a screen of approximately 5000 lines, we identified constitutive disease resistance (CDR1) encoding an apoplastic aspartic protease, the overexpression of which causes dwarfing and resistance to virulent Pseudomonas syringae. These phenotypes reflect salicylic-acid-dependent activation of micro-oxidative bursts and various defense-related genes. Antisense CDR1 plants were compromised for resistance to avirulent P. syringae and more susceptible to virulent strains than wild type. CDR1 accumulates in intercellular fluid in response to pathogen attacks. Induction of CDR1 generates a small mobile signal, and CDR1 action is blocked by the protease inhibitor pepstatin and by mutations in the protease active sites. We propose that CDR1 mediates a peptide signal system involved in the activation of inducible resistance mechanisms.
Figures



References
-
- Altschul SF, Gish W, Miller W, Myers EW, Lipman DJ (1990) Basic local alignment search tool. J Mol Biol 215: 403–410 - PubMed
-
- Alvarez ME, Pennell R, Meijer P-J, Ishikawa A, Dixon RA, Lamb C (1998) Reactive oxygen intermediates mediate a systemic signal network in the establishment of plant immunity. Cell 92: 773–784 - PubMed
-
- Aoyama T, Chua N-H (1997) A glucocorticoid-mediated transcriptional induction system in transgenic plants. Plant J 11: 605–612 - PubMed
-
- Barrett AJ (1998) Proteolytic enzymes: nomenclature and classification. In Handbook of Proteolytic Enzymes, Woessner JF (ed) London: Academic Press pp 1–20
Publication types
MeSH terms
Substances
Associated data
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
