

Two bacteria phylotypes are predominant in the Suiyo seamount hydrothermal plume
- PMID: 14766605
- PMCID: PMC348851
- DOI: 10.1128/AEM.70.2.1190-1198.2004
Two bacteria phylotypes are predominant in the Suiyo seamount hydrothermal plume
Abstract
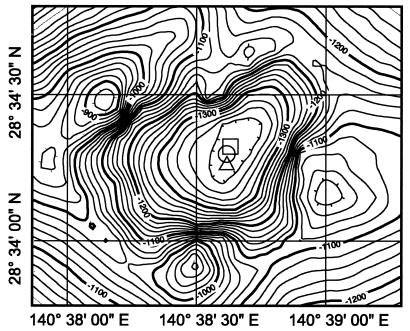
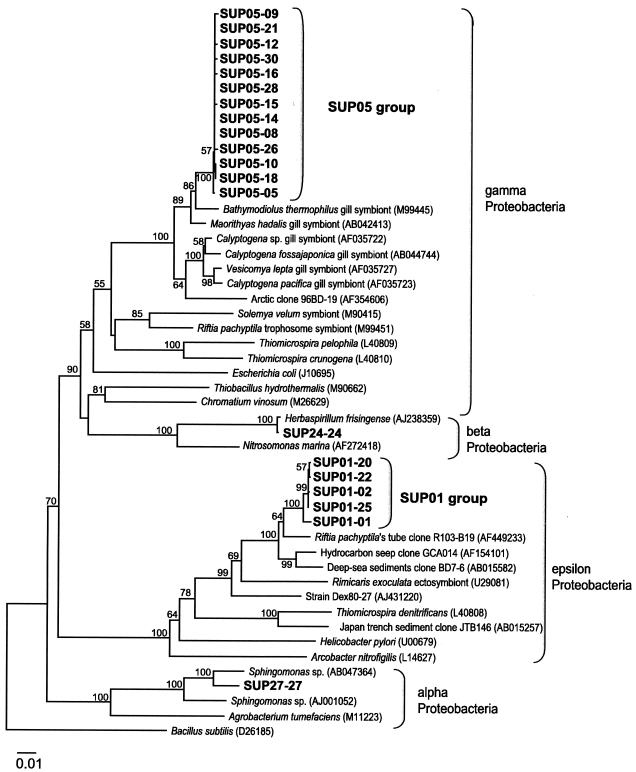
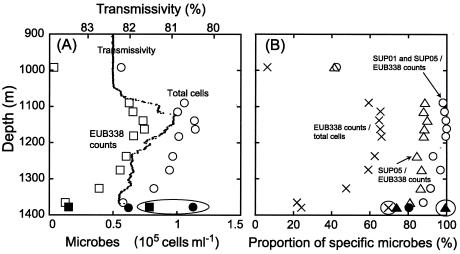
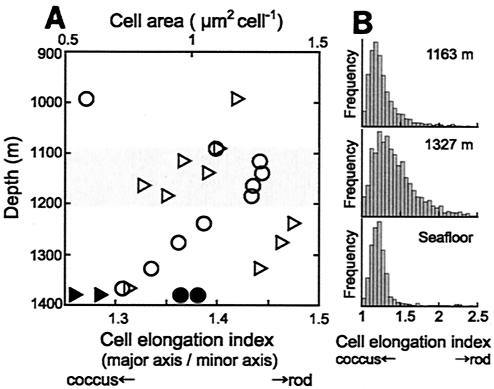
Microbial diversity and populations in a hydrothermal plume that was present inside the caldera of the Suiyo Seamount, a submarine volcano on the Izu-Bonin Arc, were investigated by performing a phylogenetic analysis of the 16S rRNA gene and by using fluorescence in situ hybridization (FISH). Corresponding to transmissivity, an indicator of turbidity, the vertical total cell count as determined by 4',6'-diamidino-2-phenylindole (DAPI) staining varied from 5.6 x 10(4) to 1.1 x 10(5) cells ml(-1), and the apparent plume layer was assessed to be at a depth of 1,050 to 1,200 m inside the caldera and to contain 1.0 x 10(5) to 1.1 x 10(5) cells ml(-1). From microbial samples collected in the plume by an in situ filtration system, the following two major phylogenetic groups, which were closely related to sulfur-oxidizing microbes, were obtained: the SUP05 group belonging to the gamma subclass of the Proteobacteria (13 of 20 clones) and the SUP01 group belonging to the epsilon subclass of the Proteobacteria (5 of 20 clones). Specific oligonucleotide probes for these groups (SUP05-187 and SUP01-63) were designed and were used with various water samples obtained from the Suiyo Seamount. In the apparent plume layer, up to 66% of the total counts of microbial cells were estimated to be Bacteria cells that hybridized to EUB338, and few cells were identified by the archaeal probe ARCH915. Almost all Bacteria cells were hard to identify with the known group-specific probes, such as ALF19, GAM42a, and CF319, while 88 to 90% of the Bacteria cells hybridized with SUP05-187 and >98% of them were considered members of the SUP05 and SUP01 populations. In a low-temperature vent fluid emitted from a bivalve-colonized mound, the SUP05 cells accounted for >99% of the Bacteria cells, suggesting that a portion of the plume cells originated on the surface of the seafloor at a depth of about 1,380 m. From further analysis of cell morphology (i.e., cell size and cell elongation index) we inferred that the SUP05 cells were active in the plume layer at a depth of 1,050 to 1,200 m compared to the activity in a near-bottom layer, while many elongated cells were found between these layers. These findings suggest that the morphology and distribution of SUP05 cells have complex relationships with hydrothermal activities and water circulation. Although growth and production rates remain to be defined, we concluded that this Suiyo Seamount caldera has functioned as a natural continuous incubator for these two phylotypes of Bacteria in an aphotic deep-sea environment.
Figures

References
-
- Brosius, J., T. J. Dull, D. D. Sleeter, and H. F. Noller. 1981. Gene organization and primary structure of a ribosomal RNA operon from Escherichia coli. J. Mol. Biol. 148:107-127. - PubMed
-
- Campbell, B. J., C. Jeanthon, J. E. Kostka, G. W. Luther III, and S. C. Cary. 2001. Growth and phylogenetic properties of novel bacteria belonging to the epsilon subdivision of the Proteobacteria enriched from Alvinella pompejana and deep-sea hydrothermal vents. Appl. Environ. Microbiol. 67:4566-4572. - PMC - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
