

B lineage-specific regulation of V(D)J recombinase activity is established in common lymphoid progenitors
- PMID: 14769852
- PMCID: PMC2211824
- DOI: 10.1084/jem.20031800
B lineage-specific regulation of V(D)J recombinase activity is established in common lymphoid progenitors
Abstract
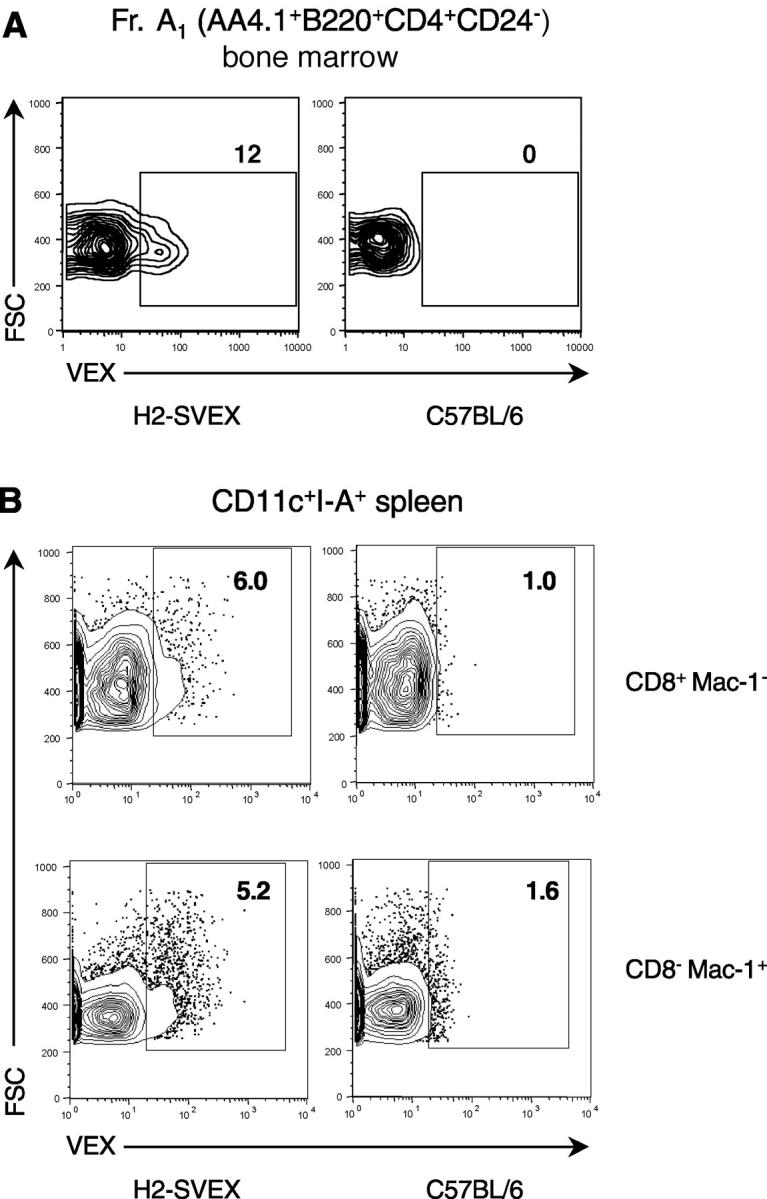
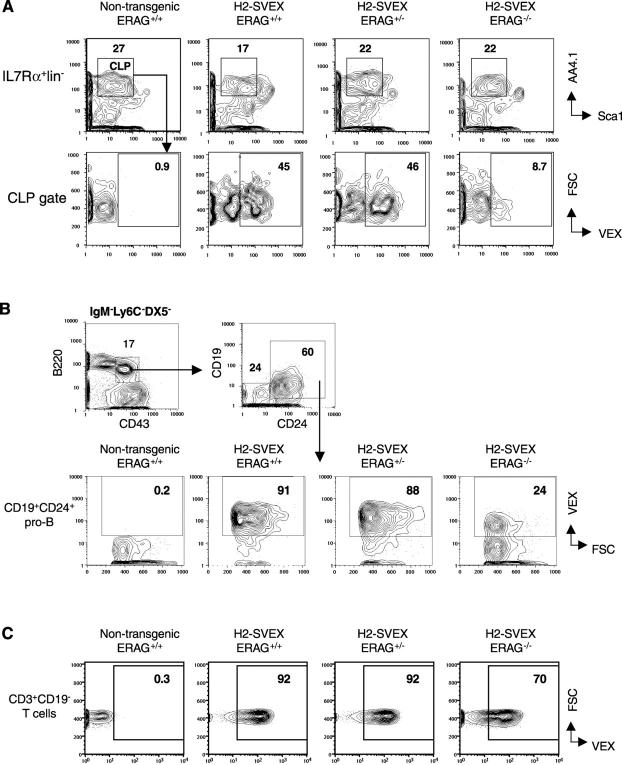
Expression of V(D)J recombinase activity in developing lymphocytes is absolutely required for initiation of V(D)J recombination at antigen receptor loci. However, little is known about when during hematopoietic development the V(D)J recombinase is first active, nor is it known what elements activate the recombinase in multipotent hematopoietic progenitors. Using mice that express a fluorescent transgenic V(D)J recombination reporter, we show that the V(D)J recombinase is active as early as common lymphoid progenitors (CLPs) but not in the upstream progenitors that retain myeloid lineage potential. Evidence of this recombinase activity is detectable in all four progeny lineages (B, T, and NK, and DC), and rag2 levels are the highest in progenitor subsets immediately downstream of the CLP. By single cell PCR, we demonstrate that V(D)J rearrangements are detectable at IgH loci in approximately 5% of splenic natural killer cells. Finally, we show that recombinase activity in CLPs is largely controlled by the Erag enhancer. As activity of the Erag enhancer is restricted to the B cell lineage, this provides the first molecular evidence for establishment of a lineage-specific transcription program in multipotent progenitors.
Figures
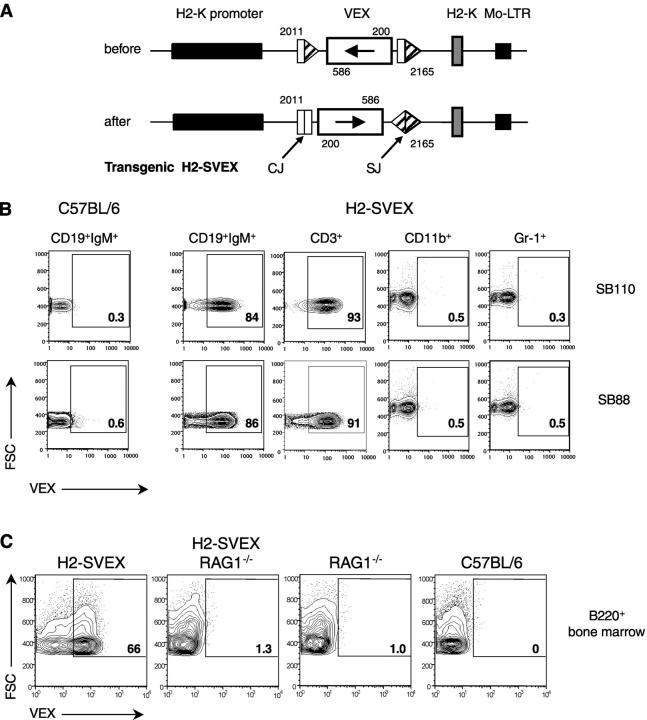
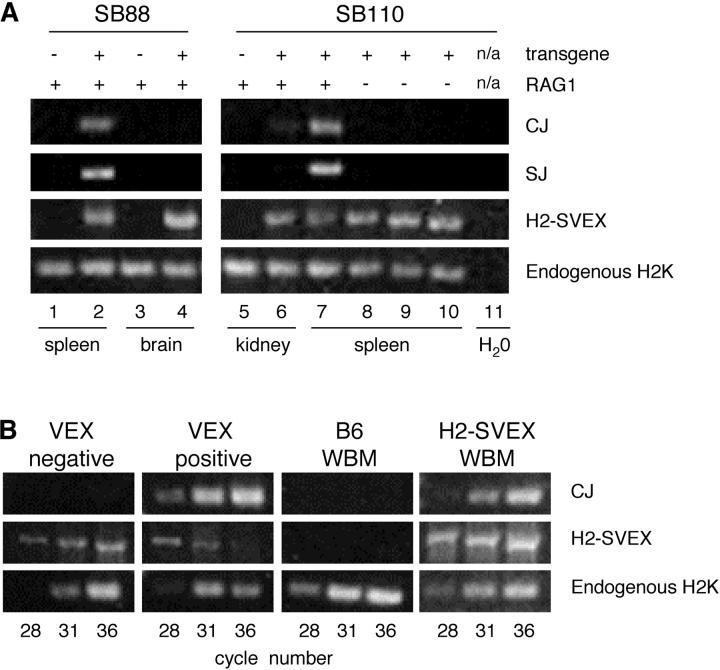
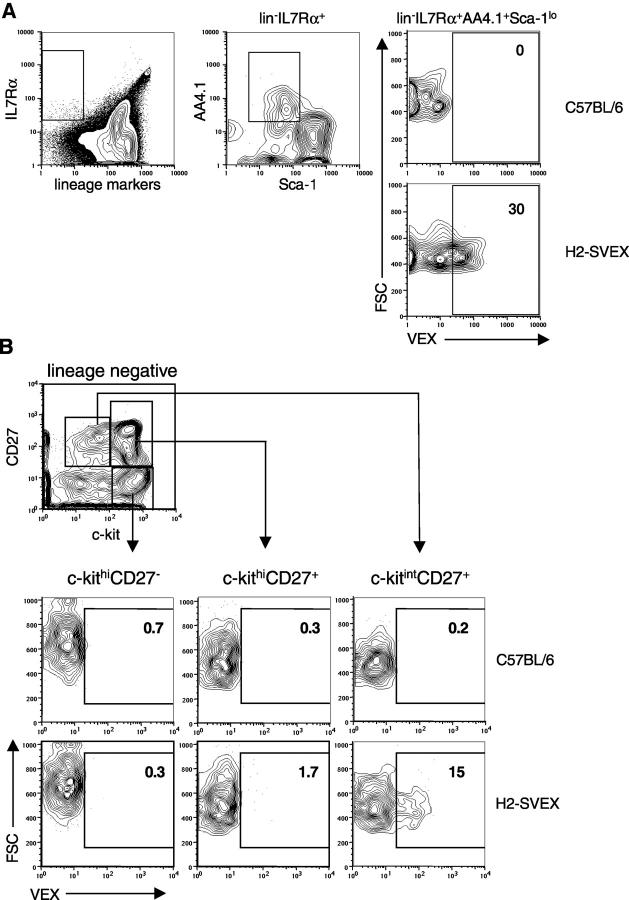
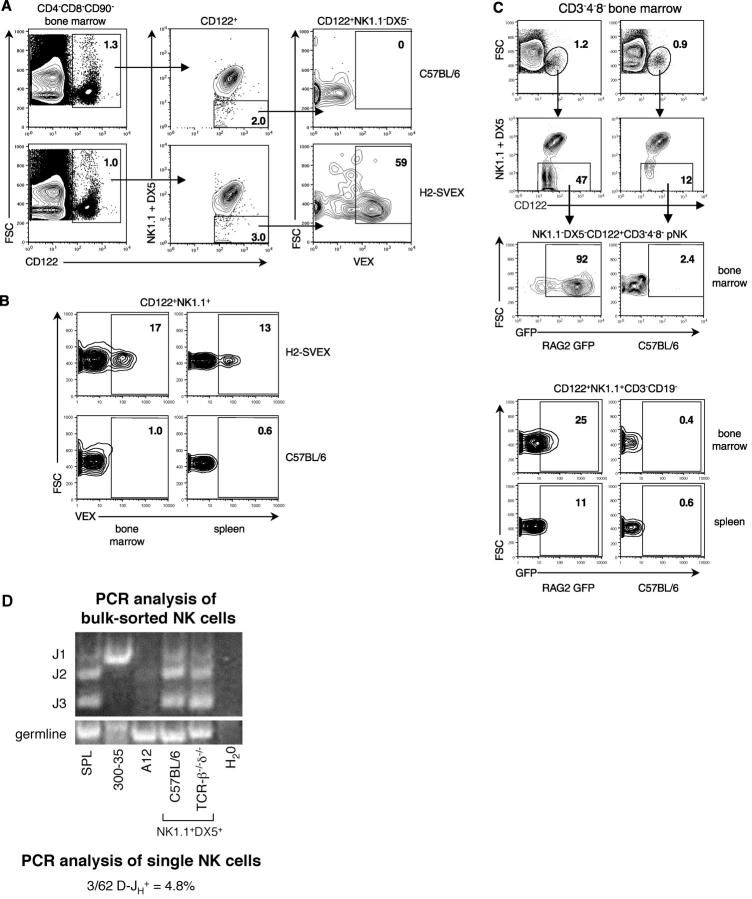
References
-
- Nutt, S.L., D. Eberhard, M. Horcher, A.G. Rolink, and M. Busslinger. 2001. Pax5 determines the identity of B cells from the beginning to the end of B-lymphopoiesis. Int. Rev. Immunol. 20:65–82. - PubMed
-
- Rothenberg, E.V., and C.J. Dionne. 2002. Lineage plasticity and commitment in T-cell development. Immunol. Rev. 187:96–115. - PubMed
-
- Mombaerts, P., J. Iacomini, R.S. Johnson, K. Herrup, S. Tonegawa, and V.E. Papaioannou. 1992. RAG-1-deficient mice have no mature B and T lymphocytes. Cell. 68:869–877. - PubMed
-
- Shinkai, Y., G. Rathbun, K.P. Lam, E.M. Oltz, V. Stewart, M. Mendelsohn, J. Charron, M. Datta, F. Young, A.M. Stall, et al. 1992. RAG-2-deficient mice lack mature lymphocytes owing to inability to initiate V(D)J rearrangement. Cell. 68:855–867. - PubMed
-
- Gao, Y., J. Chaudhuri, C. Zhu, L. Davidson, D.T. Weaver, and F.W. Alt. 1998. A targeted DNA-PKcs-null mutation reveals DNA-PK-independent functions for KU in V(D)J recombination. Immunity. 9:367–376. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Miscellaneous
