

Mimicking live flavivirus immunization with a noninfectious RNA vaccine
- PMID: 14769933
- PMCID: PMC357033
- DOI: 10.1073/pnas.0307145101
Mimicking live flavivirus immunization with a noninfectious RNA vaccine
Abstract
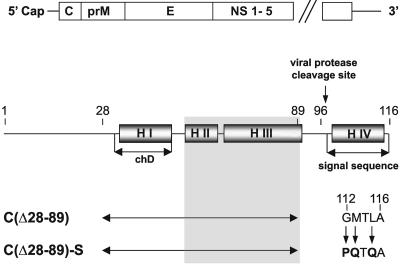
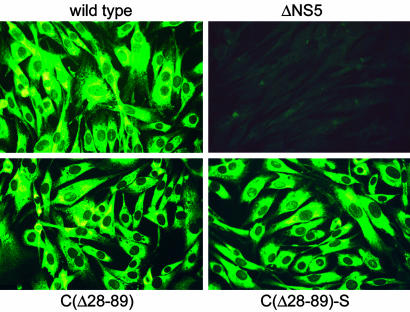
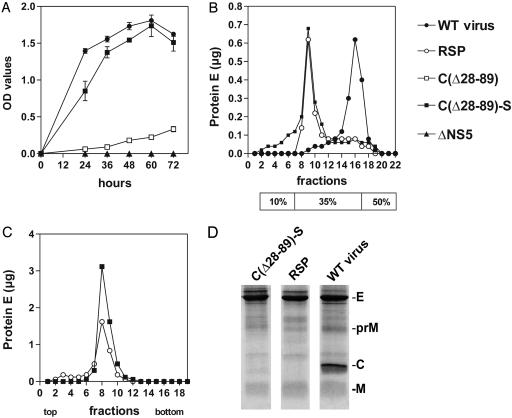
Flaviviruses are human pathogens of world-wide medical importance. They have recently received much additional attention because of their spread to new regions (such as West Nile virus to North America), highlighting their potential as newly emerging disease agents. Using tick-borne encephalitis virus, we have developed and evaluated in mice a new genetic vaccine based on self-replicating but noninfectious RNA. This RNA contains all of the necessary genetic information for establishing its replication machinery in the host cell, thus mimicking a natural infection. However, genetic modifications in the region encoding the capsid protein simultaneously prevent the assembly of infectious virus particles and promote the secretion of noninfectious subviral particles that elicit neutralizing antibodies. These characteristics demonstrate that a new generation of flavivirus vaccines can be designed that stimulate the same spectrum of innate and specific immune responses as a live vaccine but have the safety features of an inactivated vaccine.
Figures
References
-
- Lindenbach, B. D. & Rice, C. M. (2001) in Fields Virology, eds. Knipe, D. M., Howley, P. M., et al. (Lippincott Williams & Wilkins, Philadelphia), pp. 991–1041.
-
- Pugachev, K. V., Guirakhoo, F., Trent, D. W. & Monath, T. P. (2003) Int. J. Parasitol. 33, 567–582. - PubMed
-
- Murphy, B. R. & Chanock, R. M. (2001) in Fields Virology, eds. Knipe, D. M., Howley, P. M., et al. (Lippincott Williams & Wilkins, Philadelphia), pp. 435–467.
-
- Monath, T. P., Guirakhoo, F., Nichols, R., Yoksan, S., Schrader, R., Murphy, C., Blum, P., Woodward, S., McCarthy, K., Mathis, D. et al. (2003) J. Infect. Dis. 188, 1213–1230. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
