

Response of the Saccharomyces cerevisiae Mpk1 mitogen-activated protein kinase pathway to increases in internal turgor pressure caused by loss of Ppz protein phosphatases
- PMID: 14871941
- PMCID: PMC329512
- DOI: 10.1128/EC.3.1.100-107.2004
Response of the Saccharomyces cerevisiae Mpk1 mitogen-activated protein kinase pathway to increases in internal turgor pressure caused by loss of Ppz protein phosphatases
Abstract
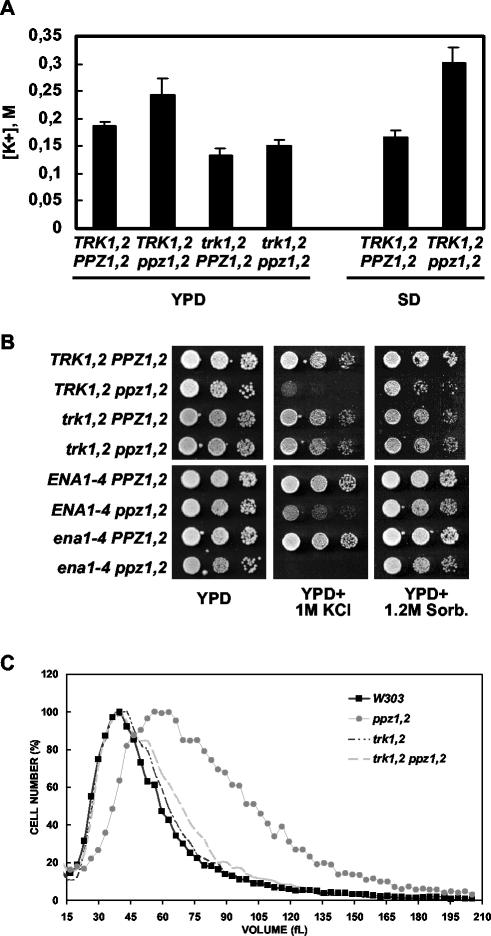
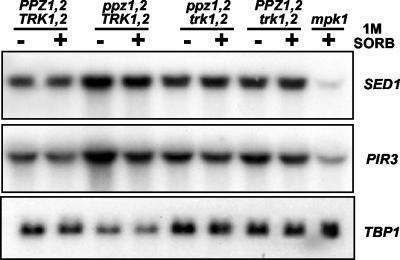
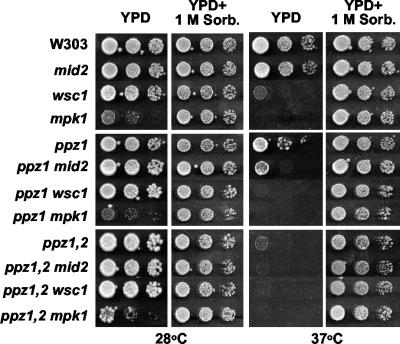
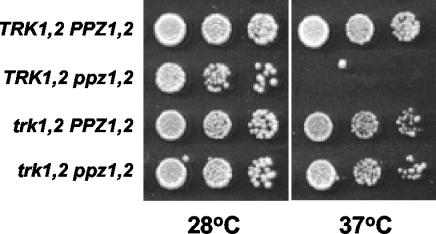
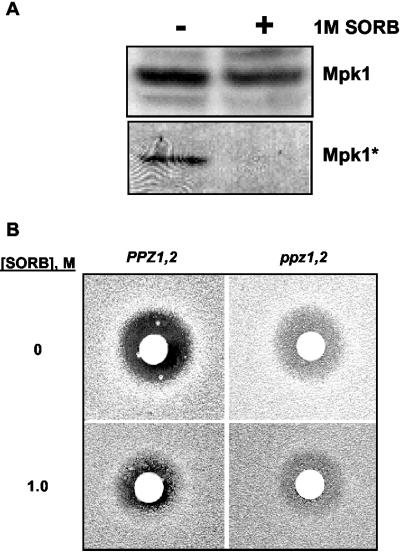
The Mpk1 pathway of Saccharomyces cerevisiae is a key determinant of cell wall integrity. A genetic link between the Mpk1 kinase and the Ppz phosphatases has been reported, but the nature of this connection was unclear. Recently, the Ppz phosphatases were shown to be regulators of K+ and pH homeostasis. Here, we demonstrate that Ppz-deficient strains display increased steady-state K+ levels and sensitivity to increased KCl concentrations. Given these observations and the fact that K+ is the major determinant of intracellular turgor pressure, we reasoned that the connection between PPZ1 and -2 and MPK1 was due to the combination of increased internal turgor pressure in Ppz-deficient strains and cell wall instability observed in strains lacking MPK1. Accordingly, the MPK1 gene was up-regulated, the Mpk1 protein was overexpressed, and the phosphorylated active form was more abundant in Ppz-deficient strains. Moreover, the expression of genes previously identified as targets of the Mpk1 pathway are also up-regulated in strains lacking PPZ1 and -2. The transcriptional and posttranslational modifications of Mpk1 were not observed when the internal K+ concentration (and thus turgor pressure) was lowered by disrupting the TRK1 and -2 K+ transporter genes or when the cell wall was stabilized by the addition of sorbitol. Moreover, we present genetic evidence showing that both the Wsc1 and Mid2 branches of the Mpk1 pathway contribute to this response. Finally, despite its role in G1/S transition, increased levels of activated Mpk1 do not appear to be responsible for the cell cycle phenotype observed in Ppz-deficient strains.
Figures


References
-
- Caro, L. H., H. Tettelin, J. H. Vossen, A. F. Ram, H. van den Ende, and F. M. Klis. 1997. In silicio identification of glycosyl-phosphatidylinositol-anchored plasma-membrane and cell wall proteins of Saccharomyces cerevisiae. Yeast 13:1477-1489. - PubMed
-
- Davenport, K. R., M. Sohaskey, Y. Kamada, D. E. Levin, and M. C. Gustin. 1995. A second osmosensing signal transduction pathway in yeast. Hypotonic shock activates the PKC1 protein kinase-regulated cell integrity pathway. J. Biol. Chem. 270:30157-30161. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
