

Impact of a disruption of a pathway delivering copper to mitochondria on Podospora anserina metabolism and life span
- PMID: 14871950
- PMCID: PMC329504
- DOI: 10.1128/EC.3.1.200-211.2004
Impact of a disruption of a pathway delivering copper to mitochondria on Podospora anserina metabolism and life span
Abstract
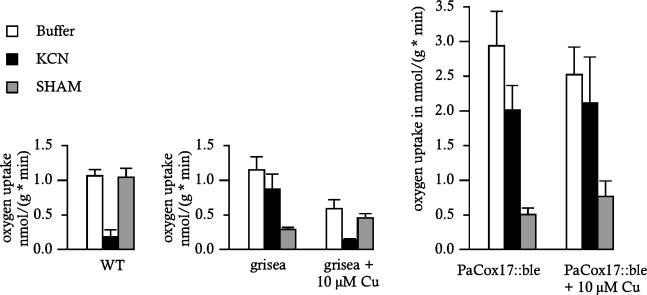
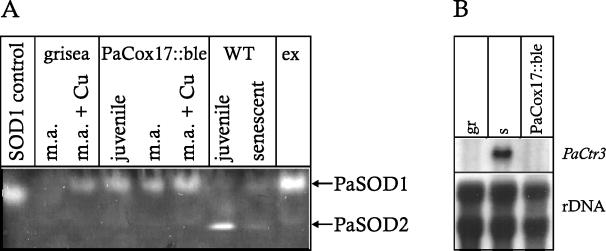
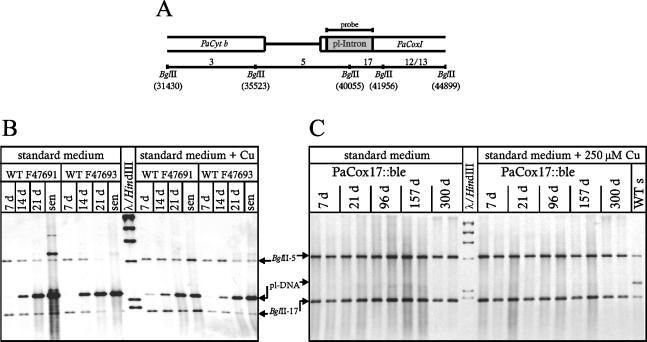
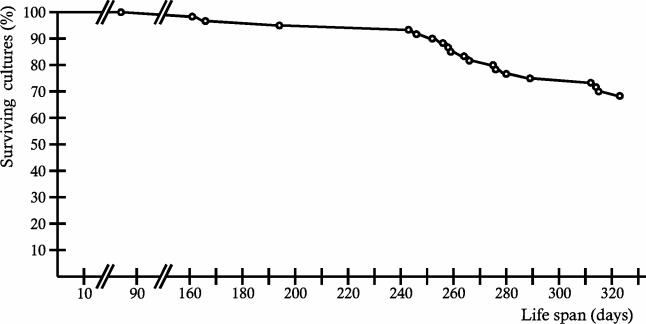
A global depletion of cellular copper as the result of a deficiency in high-affinity copper uptake was previously shown to affect the phenotype and life span of the filamentous fungus Podospora anserina. We report here the construction of a strain in which the delivery of copper to complex IV of the mitochondrial respiratory chain is affected. This strain, PaCox17::ble, is a PaCox17-null mutant that does not synthesize the molecular chaperone targeting copper to cytochrome c oxidase subunit II. PaCox17::ble is characterized by a decreased growth rate, a reduction in aerial hyphae formation, reduced female fertility, and a dramatic increase in life span. The mutant respires via a cyanide-resistant alternative pathway, displays superoxide dismutase (SOD) activity profiles significantly differing from those of the wild-type strain and is characterized by a stabilization of the mitochondrial DNA. Collectively, the presented data define individual components of a molecular network effective in life span modulation and copper as an element with a dual effect. As a cofactor of complex IV of the respiratory chain, it is indirectly involved in the generation of reactive oxygen species (ROS) and thereby plays a life span-limiting role. In contrast, Cu/Zn SOD as a ROS-scavenging enzyme lowers molecular damage and thus positively affects life span. Such considerations explain the reported differences in life span of independent mutants and spread more light on the delicate tuning of the molecular network influencing biological ageing.
Figures
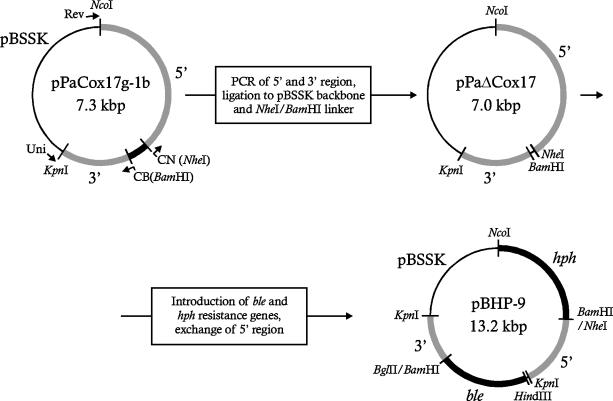
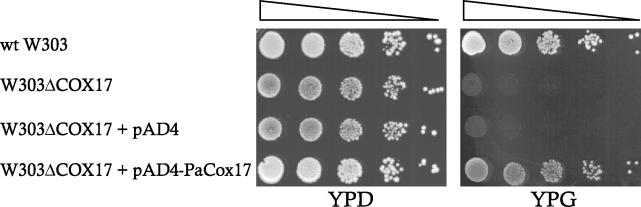
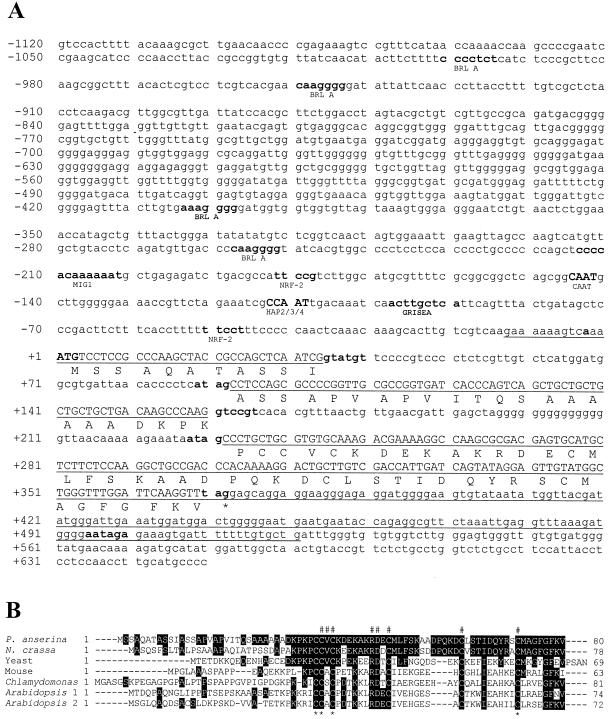
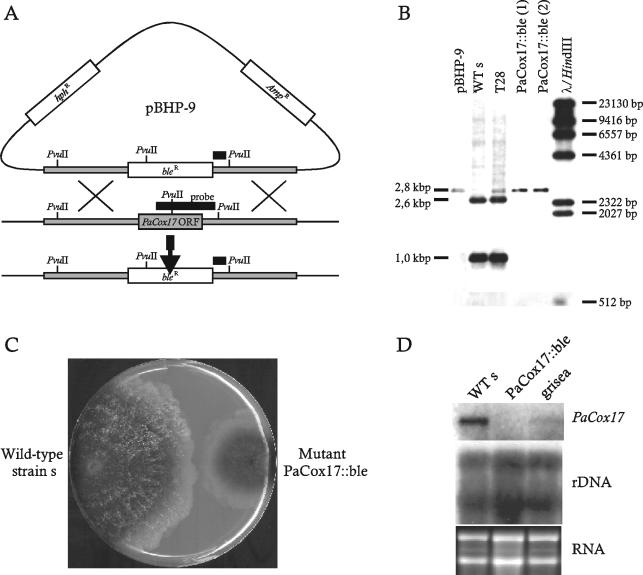

References
-
- Ballance, D. J. 1986. Sequences important for gene expression in filamentous fungi. Yeast 2:229-236. - PubMed
-
- Ballance, D. J., and G. Turner. 1986. Gene cloning in Aspergillus nidulans: isolation of the isocitrate lyase gene (acuD). Mol. Gen. Genet. 202:271-275. - PubMed
-
- Beckman, K. B., and B. N. Ames. 1998. The free radical theory of aging matures. Physiol. Rev. 78:547-581. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
