

Analysis and recognition of 5' UTR intron splice sites in human pre-mRNA
- PMID: 14960723
- PMCID: PMC373407
- DOI: 10.1093/nar/gkh273
Analysis and recognition of 5' UTR intron splice sites in human pre-mRNA
Abstract
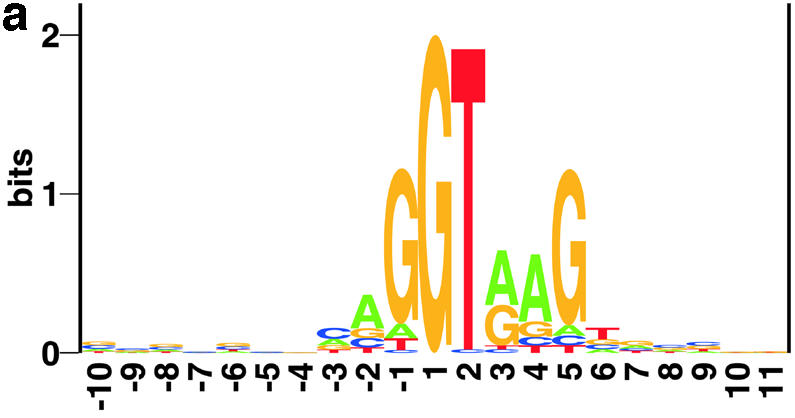
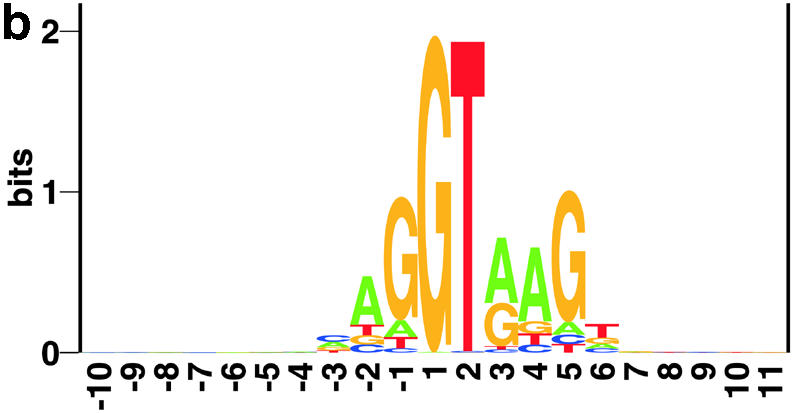
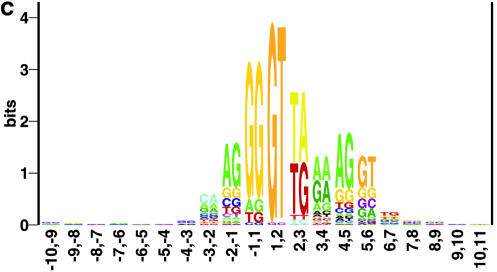
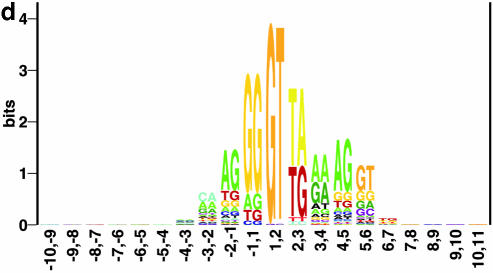
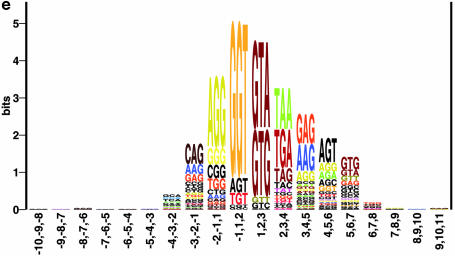
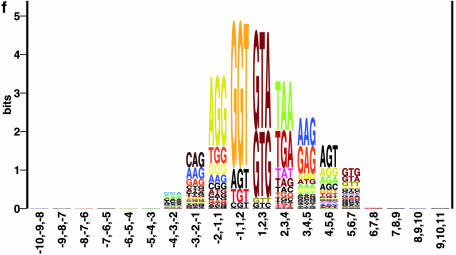
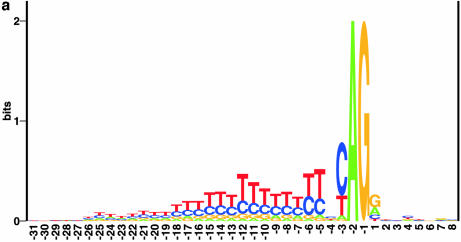
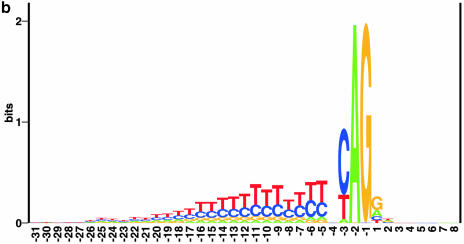
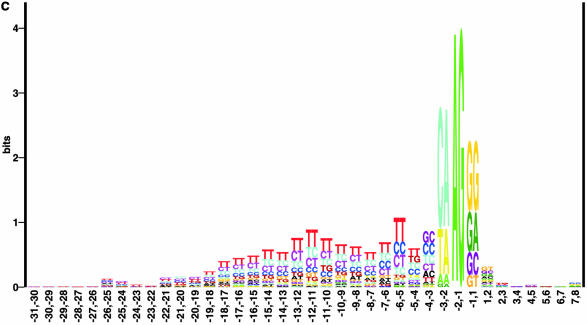
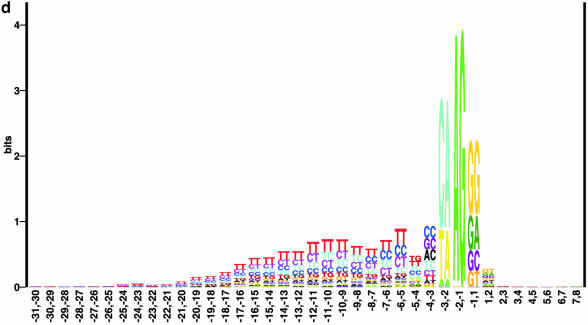
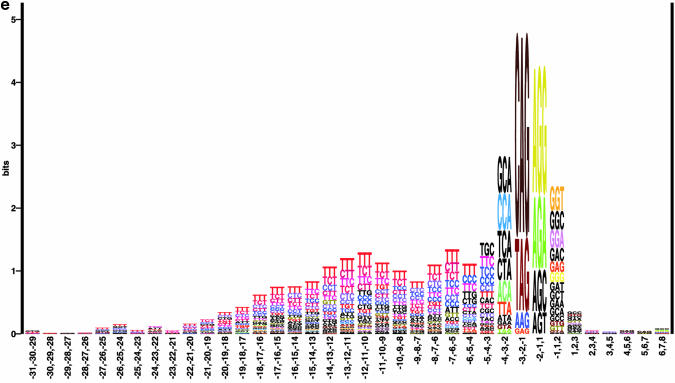
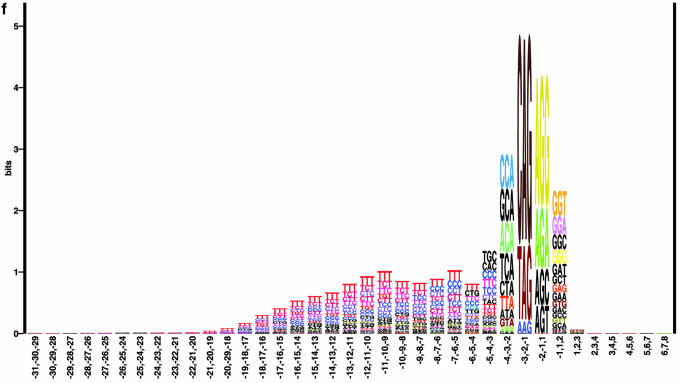
Prediction of splice sites in non-coding regions of genes is one of the most challenging aspects of gene structure recognition. We perform a rigorous analysis of such splice sites embedded in human 5' untranslated regions (UTRs), and investigate correlations between this class of splice sites and other features found in the adjacent exons and introns. By restricting the training of neural network algorithms to 'pure' UTRs (not extending partially into protein coding regions), we for the first time investigate the predictive power of the splicing signal proper, in contrast to conventional splice site prediction, which typically relies on the change in sequence at the transition from protein coding to non-coding. By doing so, the algorithms were able to pick up subtler splicing signals that were otherwise masked by 'coding' noise, thus enhancing significantly the prediction of 5' UTR splice sites. For example, the non-coding splice site predicting networks pick up compositional and positional bias in the 3' ends of non-coding exons and 5' non-coding intron ends, where cytosine and guanine are over-represented. This compositional bias at the true UTR donor sites is also visible in the synaptic weights of the neural networks trained to identify UTR donor sites. Conventional splice site prediction methods perform poorly in UTRs because the reading frame pattern is absent. The NetUTR method presented here performs 2-3-fold better compared with NetGene2 and GenScan in 5' UTRs. We also tested the 5' UTR trained method on protein coding regions, and discovered, surprisingly, that it works quite well (although it cannot compete with NetGene2). This indicates that the local splicing pattern in UTRs and coding regions is largely the same. The NetUTR method is made publicly available at www.cbs.dtu.dk/services/NetUTR.
Figures
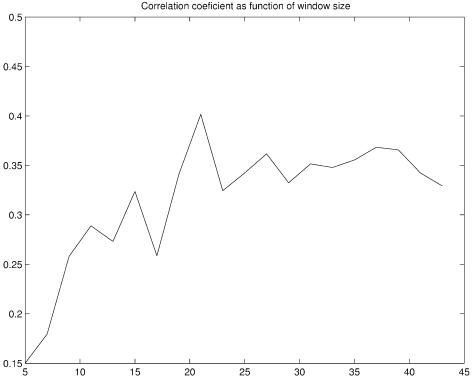

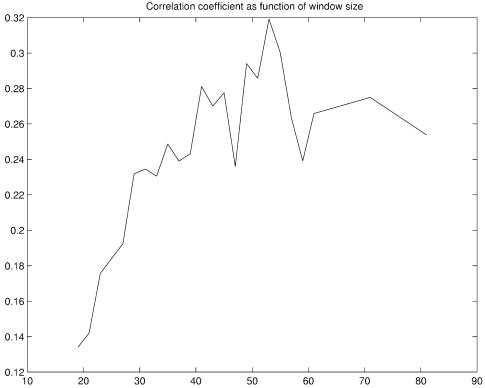
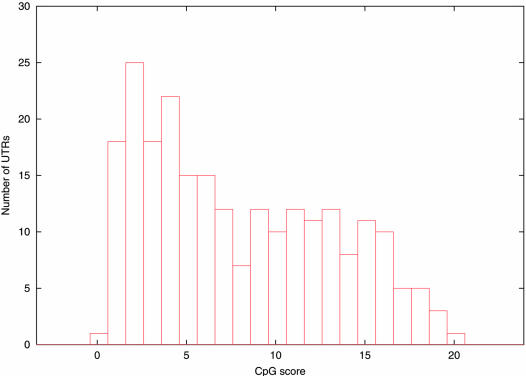
References
Publication types
MeSH terms
Substances
Associated data
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
