

Inhibition of human immunodeficiency virus type 1 replication by RNA interference directed against human transcription elongation factor P-TEFb (CDK9/CyclinT1)
- PMID: 14963154
- PMCID: PMC369228
- DOI: 10.1128/jvi.78.5.2517-2529.2004
Inhibition of human immunodeficiency virus type 1 replication by RNA interference directed against human transcription elongation factor P-TEFb (CDK9/CyclinT1)
Abstract
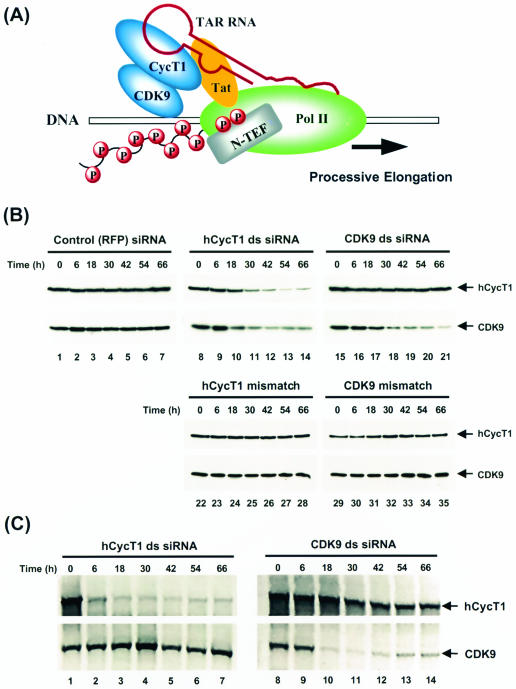
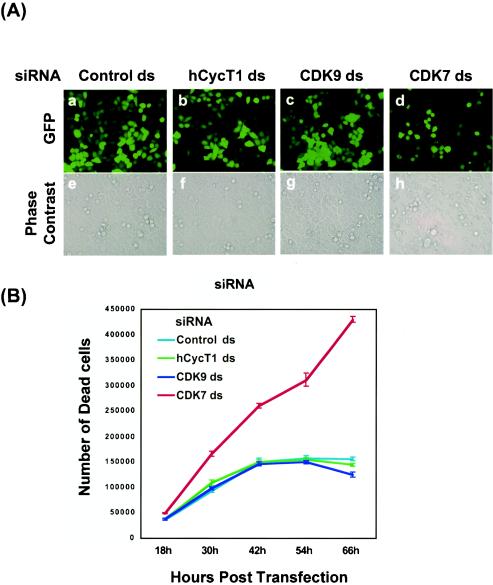
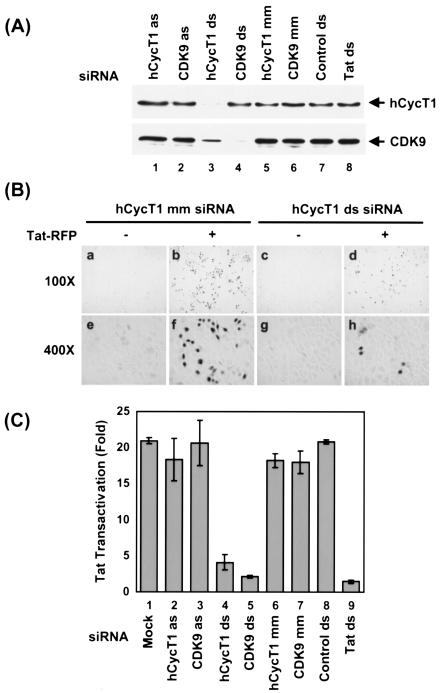
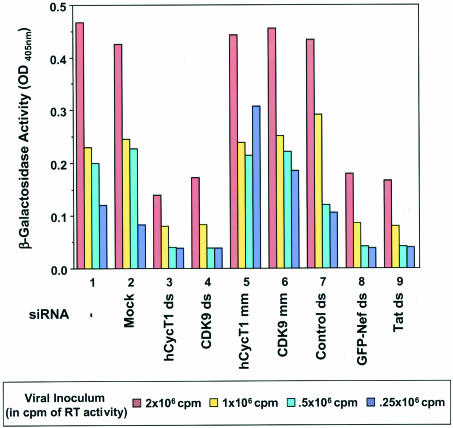
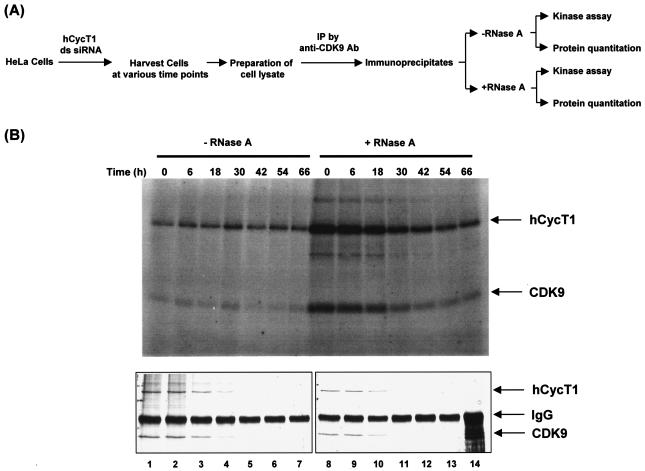
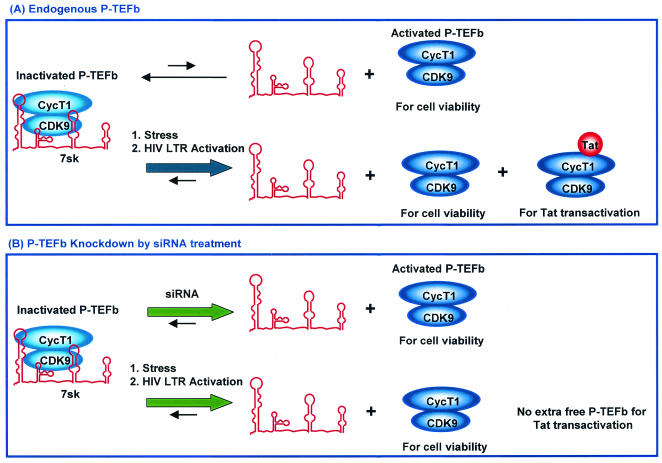
The human positive transcription elongation factor P-TEFb is composed of two subunits, cyclin T1 (hCycT1) and CDK9, and is involved in transcriptional regulation of cellular genes as well as human immunodeficiency virus type 1 (HIV-1) mRNA. Replication of HIV-1 requires the Tat protein, which activates elongation of RNA polymerase II at the HIV-1 promoter by interacting with hCycT1. To understand the cellular functions of P-TEFb and to test whether suppression of host proteins such as P-TEFb can modulate HIV infectivity without causing cellular toxicity or lethality, we used RNA interference (RNAi) to specifically knock down P-TEFb expression by degrading hCycT1 or CDK9 mRNA. RNAi-mediated gene silencing of P-TEFb in HeLa cells was not lethal and inhibited Tat transactivation and HIV-1 replication in host cells. We also found that CDK9 protein stability depended on hCycT1 protein levels, suggesting that the formation of P-TEFb CDK-cyclin complexes is required for CDK9 stability. Strikingly, P-TEFb knockdown cells showed normal P-TEFb kinase activity. Our studies suggest the existence of a dynamic equilibrium between active and inactive pools of P-TEFb in the cell and indicate that this equilibrium shifts towards the active kinase form to sustain cell viability when P-TEFb protein levels are reduced. The finding that a P-TEFb knockdown was not lethal and still showed normal P-TEFb kinase activity suggested that there is a critical threshold concentration of activated P-TEFb required for cell viability and HIV replication. These results provide new insights into the regulation of P-TEFb function and suggest the possibility that similar mechanisms for monitoring protein levels to modulate the activity of proteins may exist for the regulation of a variety of other enzymatic pathways.
Figures
Similar articles
-
Optimized chimeras between kinase-inactive mutant Cdk9 and truncated cyclin T1 proteins efficiently inhibit Tat transactivation and human immunodeficiency virus gene expression.J Virol. 2002 Nov;76(21):10873-81. doi: 10.1128/jvi.76.21.10873-10881.2002. J Virol. 2002. PMID: 12368330 Free PMC article.
-
Developmental regulators containing the I-mfa domain interact with T cyclins and Tat and modulate transcription.J Mol Biol. 2007 Mar 30;367(3):630-46. doi: 10.1016/j.jmb.2007.01.020. Epub 2007 Jan 12. J Mol Biol. 2007. PMID: 17289077 Free PMC article.
-
Regulation of TAK/P-TEFb in CD4+ T lymphocytes and macrophages.Curr HIV Res. 2003 Oct;1(4):395-404. doi: 10.2174/1570162033485159. Curr HIV Res. 2003. PMID: 15049426 Review.
-
An in vitro transcription system that recapitulates equine infectious anemia virus tat-mediated inhibition of human immunodeficiency virus type 1 Tat activity demonstrates a role for positive transcription elongation factor b and associated proteins in the mechanism of Tat activation.Virology. 2000 Sep 1;274(2):356-66. doi: 10.1006/viro.2000.0480. Virology. 2000. PMID: 10964778
-
Regulatory functions of Cdk9 and of cyclin T1 in HIV tat transactivation pathway gene expression.J Cell Biochem. 1999 Dec 1;75(3):357-68. J Cell Biochem. 1999. PMID: 10536359 Review.
Cited by
-
Antiviral RNAi: translating science towards therapeutic success.Pharm Res. 2011 Dec;28(12):2966-82. doi: 10.1007/s11095-011-0549-8. Epub 2011 Aug 9. Pharm Res. 2011. PMID: 21826573 Free PMC article. Review.
-
Cleavage and Polyadenylation Specificity Factor 6 Is Required for Efficient HIV-1 Latency Reversal.mBio. 2021 Jun 29;12(3):e0109821. doi: 10.1128/mBio.01098-21. Epub 2021 Jun 22. mBio. 2021. PMID: 34154414 Free PMC article.
-
Selective control of gene expression by CDK9 in human cells.J Cell Physiol. 2010 Jan;222(1):200-8. doi: 10.1002/jcp.21938. J Cell Physiol. 2010. PMID: 19780058 Free PMC article.
-
The emerging role of long non-coding RNAs in HIV infection.Virus Res. 2016 Jan 2;212:114-26. doi: 10.1016/j.virusres.2015.07.023. Epub 2015 Jul 26. Virus Res. 2016. PMID: 26221763 Free PMC article. Review.
-
RNA interference and antiviral therapy.World J Gastroenterol. 2007 Oct 21;13(39):5169-79. doi: 10.3748/wjg.v13.i39.5169. World J Gastroenterol. 2007. PMID: 17876887 Free PMC article. Review.
References
-
- Bai, J., J. Sui, R. Y. Zhu, A. S. Tallarico, F. Gennari, D. Zhang, and W. A. Marasco. 2003. Inhibition of Tat-mediated transactivation and HIV-1 replication by human anti-hCyclinT1 intrabodies. J. Biol. Chem. 278:1433-1442. - PubMed
-
- Chao, S. H., K. Fujinaga, J. E. Marion, R. Taube, E. A. Sausville, A. M. Senderowicz, B. M. Peterlin, and D. H. Price. 2000. Flavopiridol inhibits P-TEFb and blocks HIV-1 replication. J. Biol. Chem. 275:28345-28348. - PubMed
-
- Chiu, Y. L., and T. M. Rana. 2002. RNAi in human cells: basic structural and functional features of small interfering RNA. Mol. Cell 10:549-561. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
