

The scl +18/19 stem cell enhancer is not required for hematopoiesis: identification of a 5' bifunctional hematopoietic-endothelial enhancer bound by Fli-1 and Elf-1
- PMID: 14966269
- PMCID: PMC350551
- DOI: 10.1128/MCB.24.5.1870-1883.2004
The scl +18/19 stem cell enhancer is not required for hematopoiesis: identification of a 5' bifunctional hematopoietic-endothelial enhancer bound by Fli-1 and Elf-1
Abstract
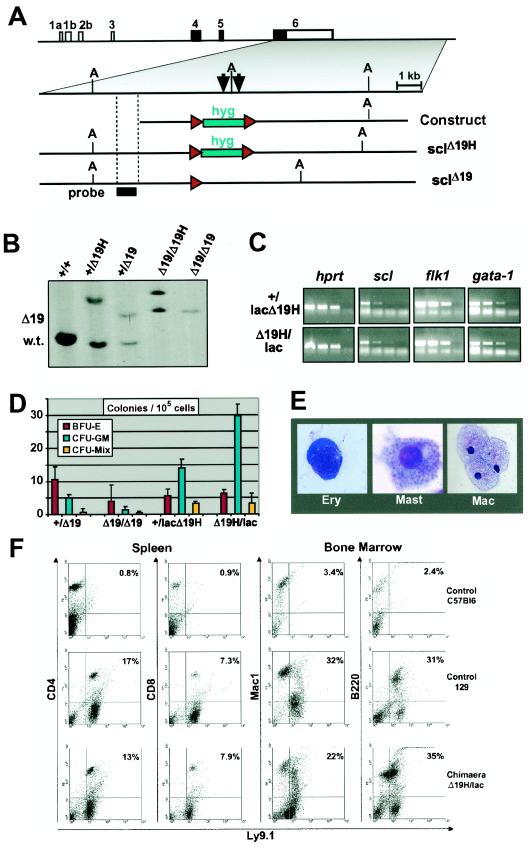
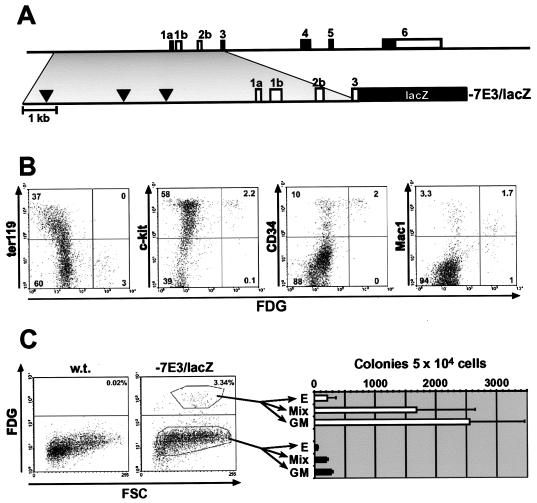
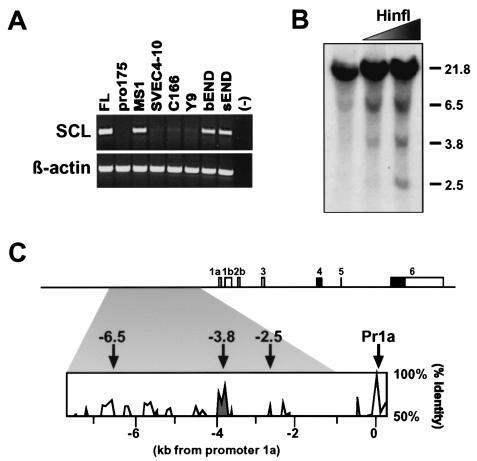
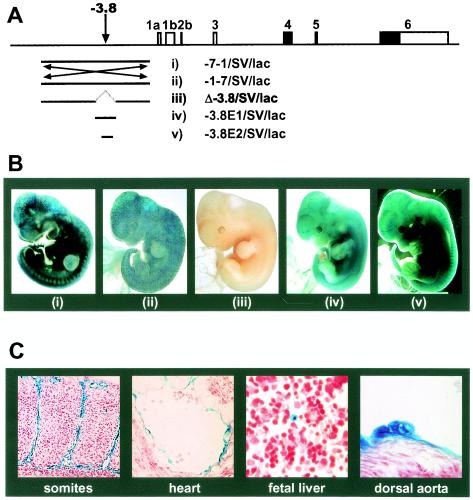
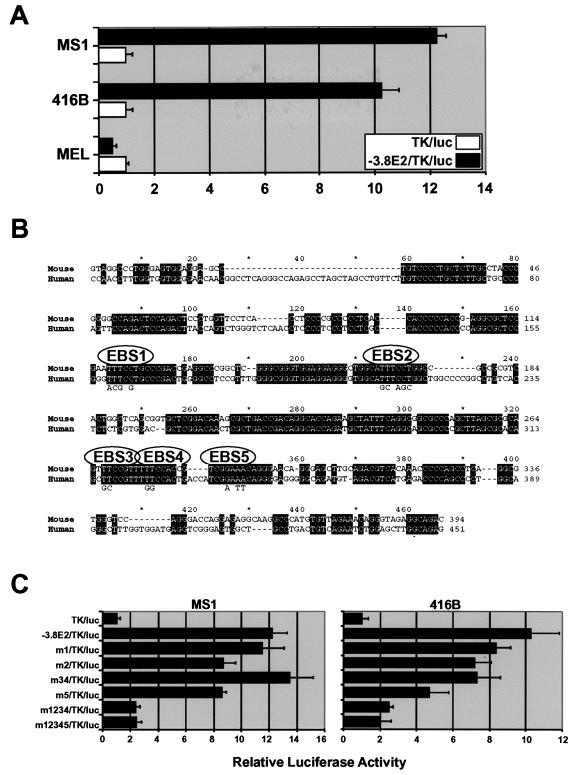
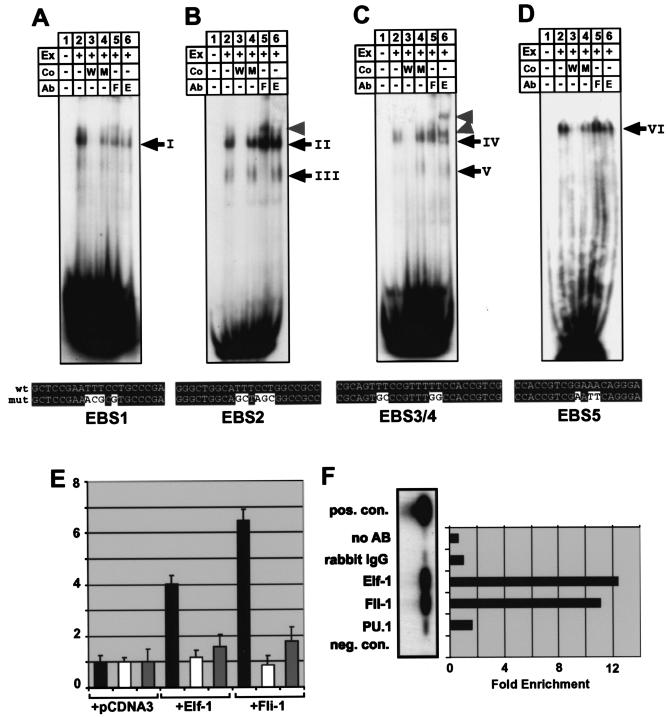
Analysis of cis-regulatory elements is central to understanding the genomic program for development. The scl/tal-1 transcription factor is essential for lineage commitment to blood cell formation and previous studies identified an scl enhancer (the +18/19 element) which was sufficient to target the vast majority of hematopoietic stem cells, together with hematopoietic progenitors and endothelium. Moreover, expression of scl under control of the +18/19 enhancer rescued blood progenitor formation in scl(-/-) embryos. However, here we demonstrate by using a knockout approach that, within the endogenous scl locus, the +18/19 enhancer is not necessary for the initiation of scl transcription or for the formation of hematopoietic cells. These results led to the identification of a bifunctional 5' enhancer (-3.8 element), which targets expression to hematopoietic progenitors and endothelium, contains conserved critical Ets sites, and is bound by Ets family transcription factors, including Fli-1 and Elf-1. These data demonstrate that two geographically distinct but functionally related enhancers regulate scl transcription in hematopoietic progenitors and endothelial cells and suggest that enhancers with dual hematopoietic-endothelial activity may represent a general strategy for regulating blood and endothelial development.
Figures
Similar articles
-
Establishing the transcriptional programme for blood: the SCL stem cell enhancer is regulated by a multiprotein complex containing Ets and GATA factors.EMBO J. 2002 Jun 17;21(12):3039-50. doi: 10.1093/emboj/cdf286. EMBO J. 2002. PMID: 12065417 Free PMC article.
-
Transcriptional regulation of the SCL locus: identification of an enhancer that targets the primitive erythroid lineage in vivo.Mol Cell Biol. 2005 Jun;25(12):5215-25. doi: 10.1128/MCB.25.12.5215-5225.2005. Mol Cell Biol. 2005. PMID: 15923636 Free PMC article.
-
The SCL +40 enhancer targets the midbrain together with primitive and definitive hematopoiesis and is regulated by SCL and GATA proteins.Mol Cell Biol. 2007 Oct;27(20):7206-19. doi: 10.1128/MCB.00931-07. Epub 2007 Aug 20. Mol Cell Biol. 2007. PMID: 17709394 Free PMC article.
-
The Hematopoietic Stem and Progenitor Cell Cistrome: GATA Factor-Dependent cis-Regulatory Mechanisms.Curr Top Dev Biol. 2016;118:45-76. doi: 10.1016/bs.ctdb.2016.01.002. Epub 2016 Feb 26. Curr Top Dev Biol. 2016. PMID: 27137654 Free PMC article. Review.
-
Concise review: Blood relatives: formation and regulation of hematopoietic stem cells by the basic helix-loop-helix transcription factors stem cell leukemia and lymphoblastic leukemia-derived sequence 1.Stem Cells. 2012 Jun;30(6):1053-8. doi: 10.1002/stem.1093. Stem Cells. 2012. PMID: 22593015 Review.
Cited by
-
Single-cell chromatin accessibility maps reveal regulatory programs driving early mouse organogenesis.Nat Cell Biol. 2020 Apr;22(4):487-497. doi: 10.1038/s41556-020-0489-9. Epub 2020 Mar 30. Nat Cell Biol. 2020. PMID: 32231307 Free PMC article.
-
Enhancer in cancer pathogenesis and treatment.Genet Mol Biol. 2023 Aug 4;46(3):e20220313. doi: 10.1590/1678-4685-GMB-2022-0313. eCollection 2023. Genet Mol Biol. 2023. PMID: 37548349 Free PMC article.
-
A novel mode of enhancer evolution: the Tal1 stem cell enhancer recruited a MIR element to specifically boost its activity.Genome Res. 2008 Sep;18(9):1422-32. doi: 10.1101/gr.077008.108. Epub 2008 Aug 7. Genome Res. 2008. PMID: 18687876 Free PMC article.
-
Deletion of the Scl +19 enhancer increases the blood stem cell compartment without affecting the formation of mature blood lineages.Exp Hematol. 2012 Jul;40(7):588-598.e1. doi: 10.1016/j.exphem.2012.02.006. Epub 2012 Mar 5. Exp Hematol. 2012. PMID: 22401818 Free PMC article.
-
Effect of transcription-factor concentrations on leukemic stem cells.Blood. 2005 Sep 1;106(5):1519-24. doi: 10.1182/blood-2005-02-0717. Epub 2005 May 24. Blood. 2005. PMID: 15914558 Free PMC article. Review.
References
-
- Akashi, K., X. He, J. Chen, H. Iwasaki, C. Niu, B. Steenhard, J. Zhang, J. Haug, and L. Li. 2003. Transcriptional accessibility for genes of multiple tissues and hematopoietic lineages is hierarchically controlled during early hematopoiesis. Blood 101:383-389. - PubMed
-
- Anguita, E., J. A. Sharpe, J. A. Sloane-Stanley, C. Tufarelli, D. R. Higgs, and W. G. Wood. 2002. Deletion of the mouse alpha-globin regulatory element (HS-26) has an unexpectedly mild phenotype. Blood 100:3450-3456. - PubMed
-
- Baltzinger, M., A. M. Mager-Heckel, and P. Remy. 1999. Xl erg: expression pattern and overexpression during development plead for a role in endothelial cell differentiation. Dev. Dyn. 216:420-433. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous