

Capacitance measurements of exocytosis in mouse pancreatic alpha-, beta- and delta-cells within intact islets of Langerhans
- PMID: 14966302
- PMCID: PMC1664984
- DOI: 10.1113/jphysiol.2003.059675
Capacitance measurements of exocytosis in mouse pancreatic alpha-, beta- and delta-cells within intact islets of Langerhans
Abstract
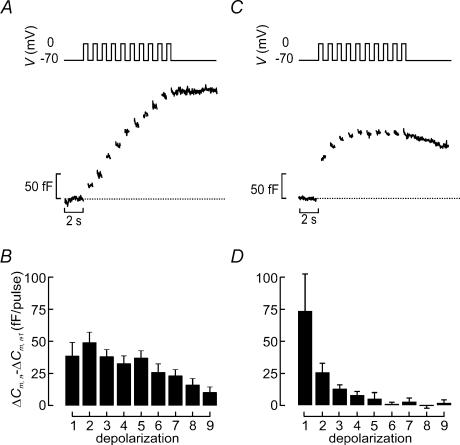
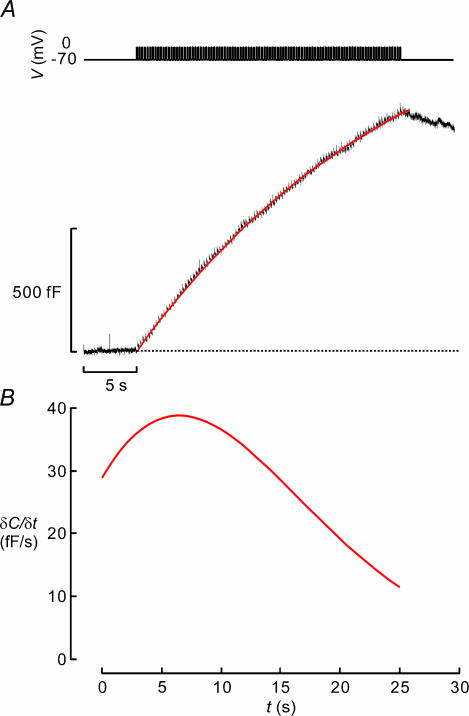
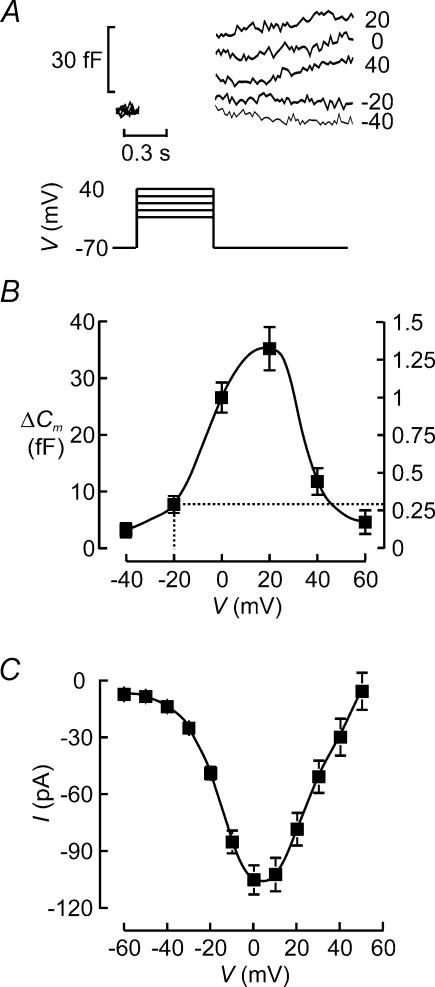
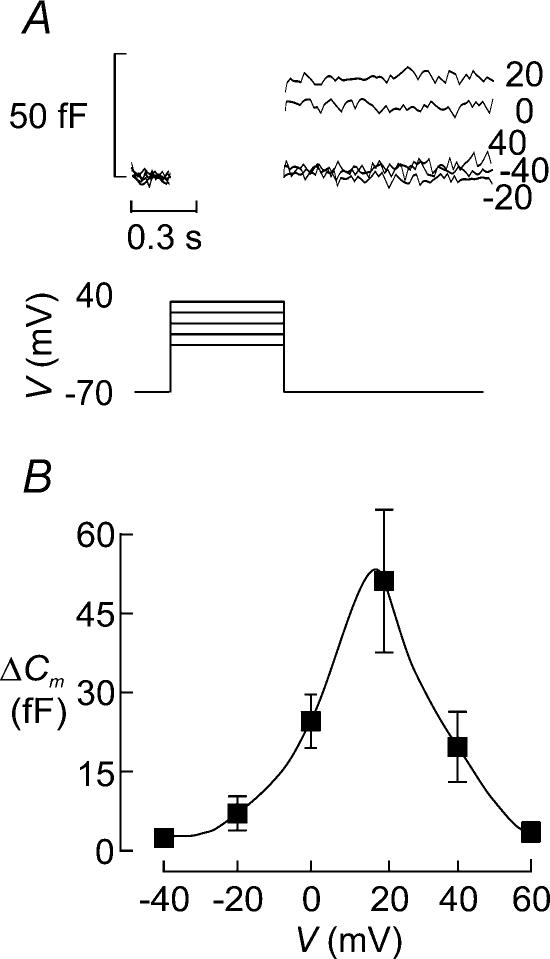
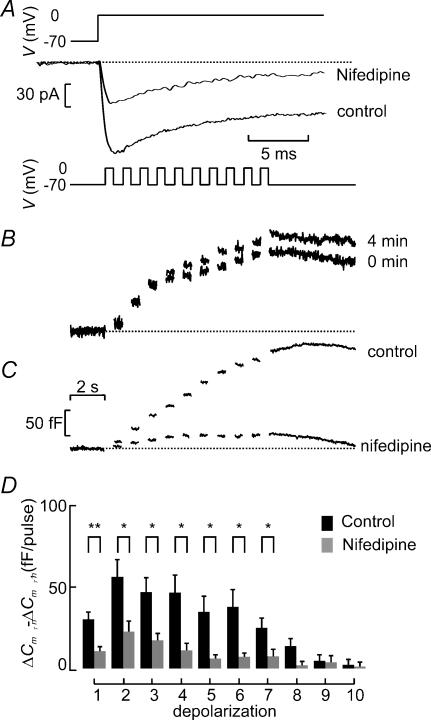
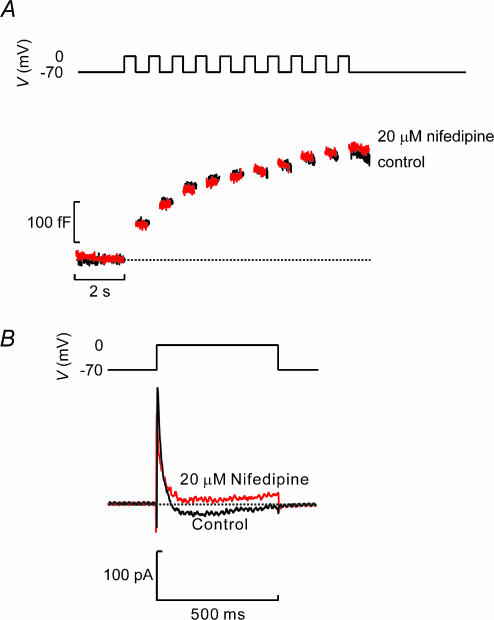
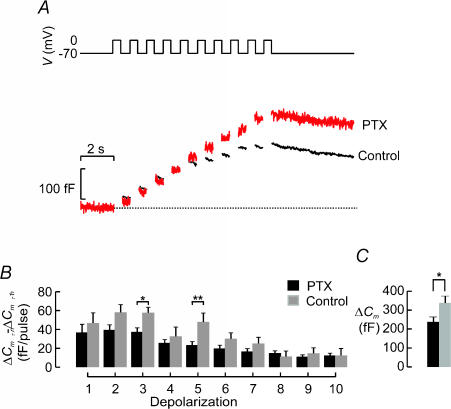
Capacitance measurements of exocytosis were applied to functionally identified alpha-, beta- and delta-cells in intact mouse pancreatic islets. The maximum rate of capacitance increase in beta-cells during a depolarization to 0 mV was equivalent to 14 granules s(-1), <5% of that observed in isolated beta-cells. Beta-cell secretion exhibited bell-shaped voltage dependence and peaked at +20 mV. At physiological membrane potentials (up to approximately -20 mV) the maximum rate of release was approximately 4 granules s(-1). Both exocytosis (measured by capacitance measurements) and insulin release (detected by radioimmunoassay) were strongly inhibited by the L-type Ca(2+) channel blocker nifedipine (25 microm) but only marginally (<20%) affected by the R-type Ca(2+) channel blocker SNX482 (100 nm). Exocytosis in the glucagon-producing alpha-cells peaked at +20 mV. The capacitance increases elicited by pulses to 0 mV exhibited biphasic kinetics and consisted of an initial transient (150 granules s(-1)) and a sustained late component (30 granules s(-1)). Whereas addition of the N-type Ca(2+) channel blocker omega-conotoxin GVIA (0.1 microm) inhibited glucagon secretion measured in the presence of 1 mm glucose to the same extent as an elevation of glucose to 20 mm, the L-type Ca(2+) channel blocker nifedipine (25 microm) had no effect. Thus, glucagon release during hyperglycaemic conditions depends principally on Ca(2+)-influx through N-type rather than L-type Ca(2+) channels. Exocytosis in the somatostatin-secreting delta-cells likewise exhibited two kinetically separable phases of capacitance increase and consisted of an early rapid (600 granules s(-1)) component followed by a sustained slower (60 granules s(-1)) component. We conclude that (1) capacitance measurements in intact pancreatic islets are feasible; (2) exocytosis measured in beta-cells in situ is significantly slower than that of isolated cells; and (3) the different types of islet cells exhibit distinct exocytotic features.
Figures
References
-
- Ammala C, Ashcroft FM, Rorsman P. Calcium-independent potentiation of insulin release by cyclic AMP in single β-cells. Nature. 1993a;363:356–358. - PubMed
-
- Ashcroft FM, Rorsman P. Elecrophysiology of pancreatic islet cells. In: Scherübl HH, Hescheler J, editors. The Electrophysiology of Neuroendocrine Cells. Boca Raton, FL, USA: CRC Press; 1995. pp. 207–243.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous