

Effects of pH on vascular tone in rabbit basilar arteries
- PMID: 14966340
- PMCID: PMC2822262
- DOI: 10.3346/jkms.2004.19.1.42
Effects of pH on vascular tone in rabbit basilar arteries
Abstract
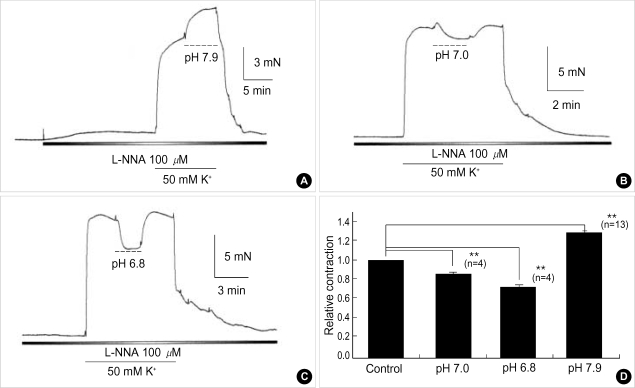
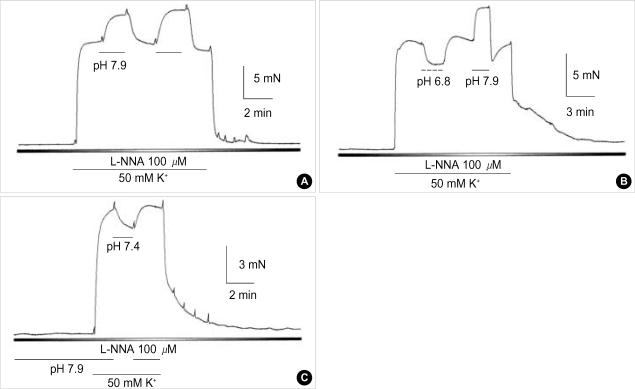
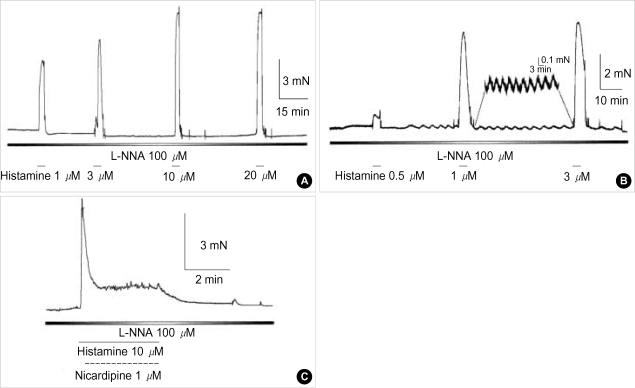
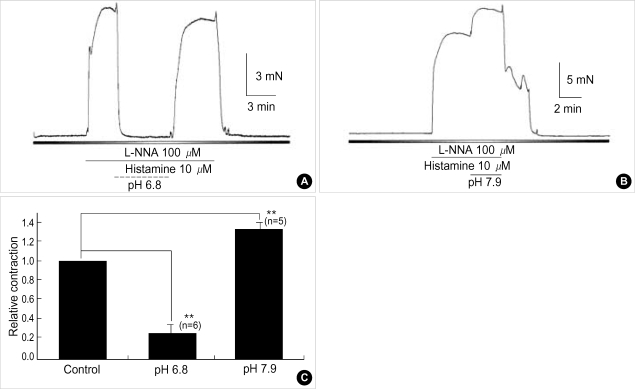
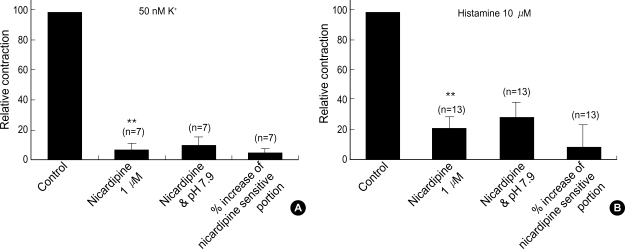
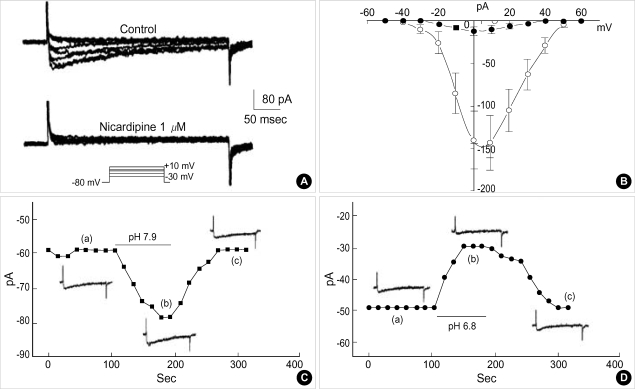
Effects of pH on vascular tone and L-type Ca2+ channels were investigated using Mulvany myograph and voltage-clamp technique in rabbit basilar arteries. In rabbit basilar arteries, high K+ produced tonic contractions by 11+/-0.6 mN (mean+/-S.E.,n=19). When extracellular pH (pHo) was changed from control 7.4 to 7.9 ([alkalosis]o), K+-induced contraction was increased to 128+/-2.1% of the control (n=13). However, K+-induced contraction was decreased to 73+/-1.3% of the control at pHo 6.8 ([acidosis] o, n=4). Histamine (10 microM) also produced tonic contraction by 11+/-0.6 mN (n=17), which was blocked by post-application of nicardipine (1 microM). [alkalosis]o and [acidosis]o increased or decreased histamine-induced contraction to 134+/-5.7% and 27+/-7.6% of the control (n=4, 6). Since high K+- and histamine-induced tonic contractions were affected by nicardipine and pHo, the effect of pHo on voltage-dependent L-type Ca2+ channel (VDCCL) was studied. VDCCL was modulated by pHo: the peak value of Ca2+ channel current (IBa) at a holding of 0 mV decreased in [acidosis]o by 41+/-8.8%, whereas that increased in [alkalosis]o by 35+/-2.1% (n=3). These results suggested that the external pH regulates vascular tone partly via the modulation of VDCC in rabbit basilar arteries.
Figures
Similar articles
-
The selective potentiation of noradrenaline-induced tone by Bay K 8644 in the rabbit basilar artery.J Cereb Blood Flow Metab. 1989 Dec;9(6):759-64. doi: 10.1038/jcbfm.1989.109. J Cereb Blood Flow Metab. 1989. PMID: 2479650
-
Cerebrovascular selectivity and vasospasmolytic action of the novel calcium antagonist (+/-)-(E)-1-(3-fluoro-6, 11-dihydrodibenz[b,e]oxepin-11-yl)-4-(3-phenyl-2-propenyl)-piperazine dimaleate in isolated cerebral arteries of the rabbit and dog.Arzneimittelforschung. 1997 Apr;47(4):339-46. Arzneimittelforschung. 1997. PMID: 9150852
-
Effects of external pH on ionic currents in smooth muscle cells from the basilar artery of the guinea pig.Circ Res. 1992 Jul;71(1):201-9. doi: 10.1161/01.res.71.1.201. Circ Res. 1992. PMID: 1606662
-
Extracellular acidosis contracts coronary but neither renal nor mesenteric artery via modulation of H+,K+-ATPase, voltage-gated K+ channels and L-type Ca2+ channels.Exp Physiol. 2014 Jul;99(7):995-1006. doi: 10.1113/expphysiol.2014.078634. Epub 2014 May 16. Exp Physiol. 2014. PMID: 24928954
-
Influence of acid-base changes on carbachol- and potassium-induced contractions of taenia coli of the rabbit.Acta Physiol Scand. 1981 Jan;111(1):59-68. doi: 10.1111/j.1748-1716.1981.tb06705.x. Acta Physiol Scand. 1981. PMID: 6784447
Cited by
-
pCO(2) and pH regulation of cerebral blood flow.Front Physiol. 2012 Sep 14;3:365. doi: 10.3389/fphys.2012.00365. eCollection 2012. Front Physiol. 2012. PMID: 23049512 Free PMC article.
-
Neuroimaging of vascular reserve in patients with cerebrovascular diseases.Neuroimage. 2019 Feb 15;187:192-208. doi: 10.1016/j.neuroimage.2017.10.015. Epub 2017 Oct 12. Neuroimage. 2019. PMID: 29031532 Free PMC article. Review.
-
Characterizing the protective vasodilatory effects of hypobaric hypoxia on the neurovascular coupling response.J Cereb Blood Flow Metab. 2025 Jul;45(7):1293-1309. doi: 10.1177/0271678X251322348. Epub 2025 Mar 13. J Cereb Blood Flow Metab. 2025. PMID: 40079563 Free PMC article.
References
-
- Tian R, Vogel P, Lassen NA, Mulvany MJ, Andreasen F, Aalkjaer C. Role of extracellular and intracellular acidosis for hypercapnia-induced inhibition of tension of isolated rat cerebral arteries. Circ Res. 1995;76:269–275. - PubMed
-
- Kontos HA. Regulation of the cerebral circulation. Annu Rev Physiol. 1981;43:397–407. - PubMed
-
- Toda N, Hatano Y, Mori K. Mechanisms underlying response to hypercapnia and bicarbonate of isolated dog cerebral arteries. Am J Physiol. 1989;257:H141–H146. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
