

pRb inactivation in mammary cells reveals common mechanisms for tumor initiation and progression in divergent epithelia
- PMID: 14966529
- PMCID: PMC340938
- DOI: 10.1371/journal.pbio.0020022
pRb inactivation in mammary cells reveals common mechanisms for tumor initiation and progression in divergent epithelia
Abstract
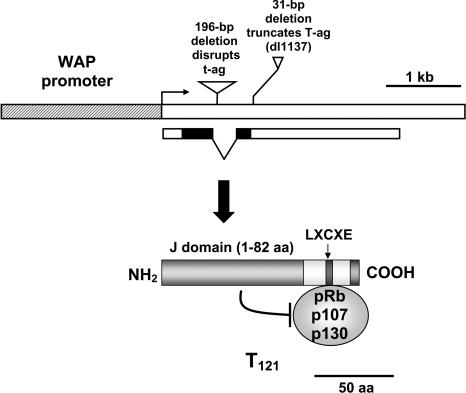
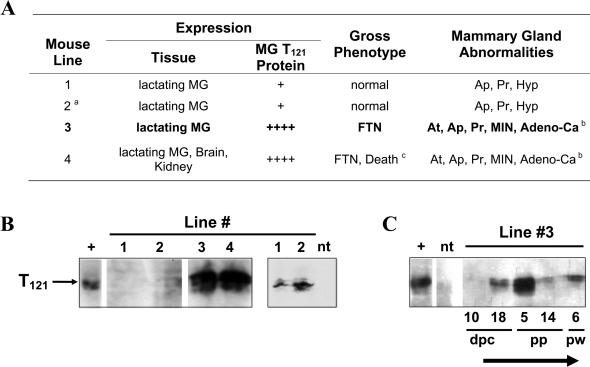
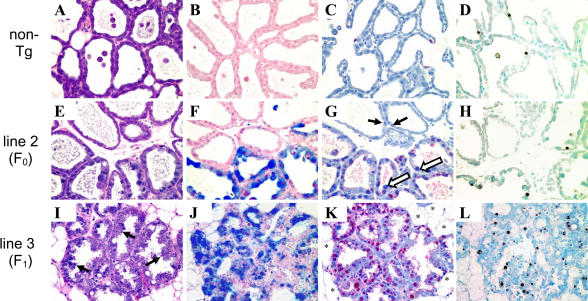
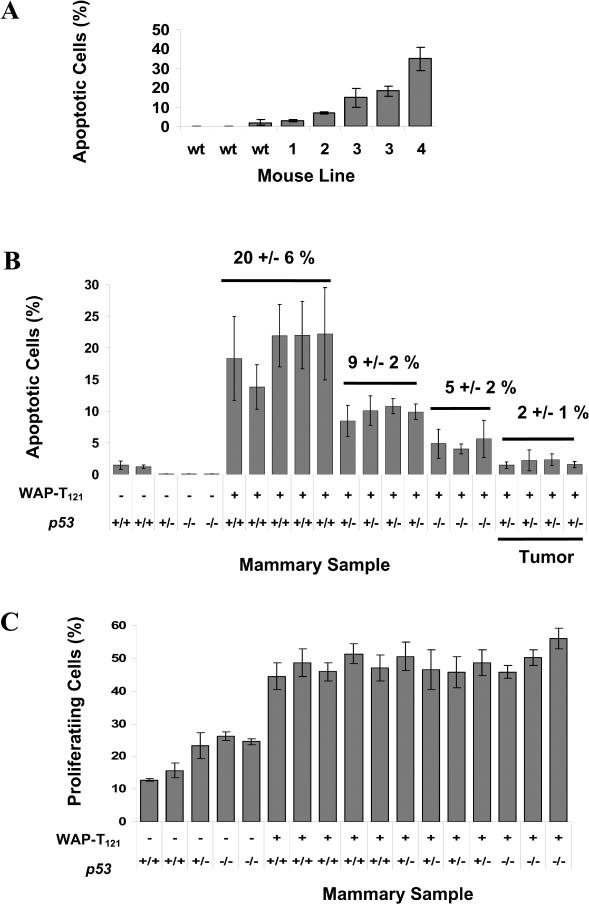
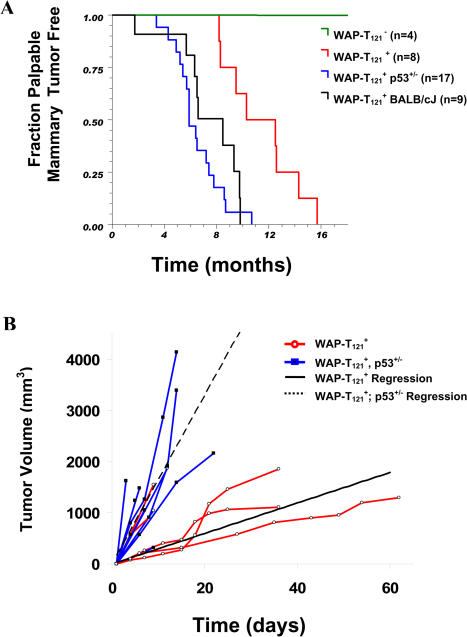
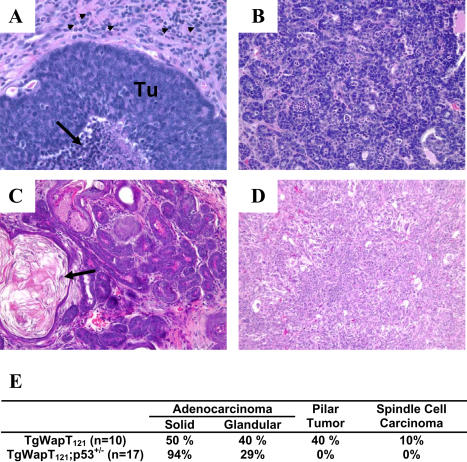
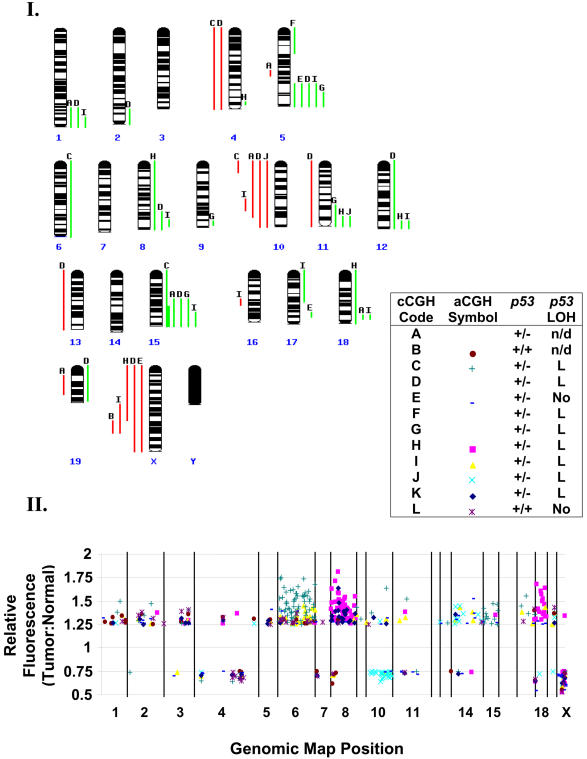
Retinoblastoma 1 (pRb) and the related pocket proteins, retinoblastoma-like 1 (p107) and retinoblastoma-like 2 (p130) (pRb(f), collectively), play a pivotal role in regulating eukaryotic cell cycle progression, apoptosis, and terminal differentiation. While aberrations in the pRb-signaling pathway are common in human cancers, the consequence of pRb(f) loss in the mammary gland has not been directly assayed in vivo. We reported previously that inactivating these critical cell cycle regulators in divergent cell types, either brain epithelium or astrocytes, abrogates the cell cycle restriction point, leading to increased cell proliferation and apoptosis, and predisposing to cancer. Here we report that mouse mammary epithelium is similar in its requirements for pRb(f) function; Rb(f) inactivation by T(121), a fragment of SV40 T antigen that binds to and inactivates pRb(f) proteins, increases proliferation and apoptosis. Mammary adenocarcinomas form within 16 mo. Most apoptosis is regulated by p53, which has no impact on proliferation, and heterozygosity for a p53 null allele significantly shortens tumor latency. Most tumors in p53 heterozygous mice undergo loss of the wild-type p53 allele. We show that the mechanism of p53 loss of heterozygosity is not simply the consequence of Chromosome 11 aneuploidy and further that chromosomal instability subsequent to p53 loss is minimal. The mechanisms for pRb and p53 tumor suppression in the epithelia of two distinct tissues, mammary gland and brain, are indistinguishable. Further, this study has produced a highly penetrant breast cancer model based on aberrations commonly observed in the human disease.
Conflict of interest statement
The authors have declared that no conflicts of interest exist.
Figures
References
-
- Brehm A, Miska EA, McCance DJ, Reid JL, Bannister AJ, et al. Retinoblastoma protein recruits histone deacetylase to repress transcription. Nature. 1998;391:597–601. - PubMed
-
- Buckley MF, Sweeney KJ, Hamilton JA, Sini RL, Manning DL, et al. Expression and amplification of cyclin genes in human breast cancer. Oncogene. 1993;8:2127–2133. - PubMed
-
- Chen J, Tobin G, Pipas JM, Van Dyke TA. T antigen mutant activities in transgenic mice: Roles of p53 and pRb-binding in tumorigenesis of the choroid plexus. Oncogene. 1992;7:1167–1175. - PubMed
-
- DeCaprio JA, Ludlow JW, Lynch D, Furukawa Y, Griffin J, et al. The product of the retinoblastoma susceptibility gene has properties of a cell-cycle regulatory element. Cell. 1989;58:1085–1095. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
