

Targeted ablation of Fgf23 demonstrates an essential physiological role of FGF23 in phosphate and vitamin D metabolism
- PMID: 14966565
- PMCID: PMC338262
- DOI: 10.1172/JCI19081
Targeted ablation of Fgf23 demonstrates an essential physiological role of FGF23 in phosphate and vitamin D metabolism
Abstract
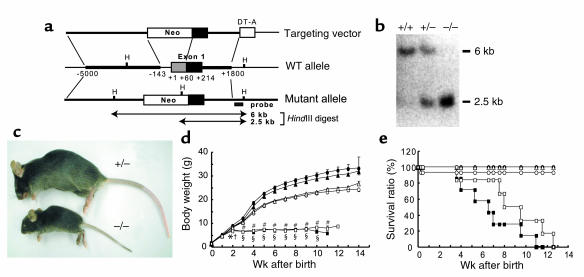
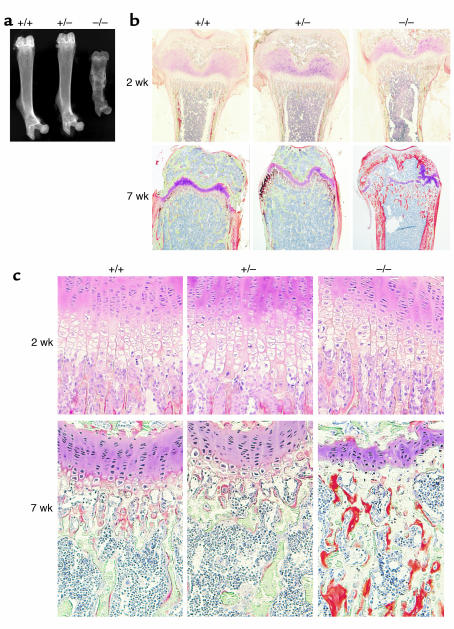
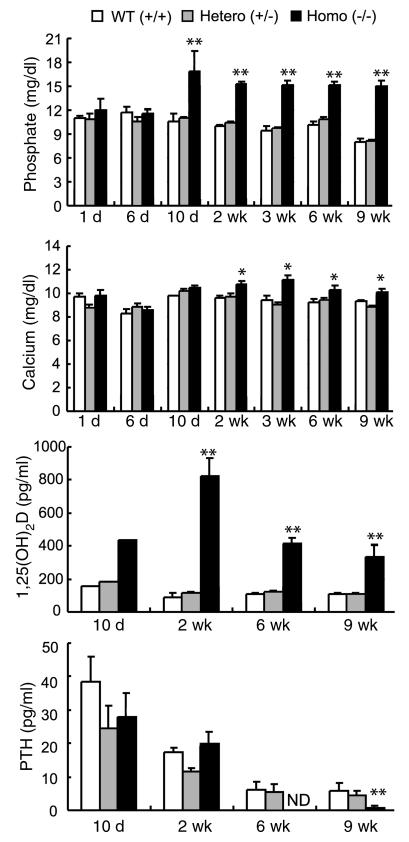
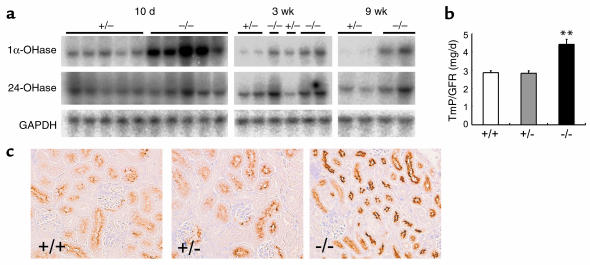
Inorganic phosphate is essential for ECM mineralization and also as a constituent of important molecules in cellular metabolism. Investigations of several hypophosphatemic diseases indicated that a hormone-like molecule probably regulates serum phosphate concentration. FGF23 has recently been recognized as playing important pathophysiological roles in several hypophosphatemic diseases. We present here the evidence that FGF23 is a physiological regulator of serum phosphate and 1,25-dihydroxyvitamin D (1,25[OH]2D) by generating FGF23-null mice. Disruption of the Fgf23 gene did not result in embryonic lethality, although homozygous mice showed severe growth retardation with abnormal bone phenotype and markedly short life span. The Fgf23(-/-) mice displayed significantly high serum phosphate with increased renal phosphate reabsorption. They also showed an elevation in serum 1,25(OH)2D that was due to the enhanced expression of renal 25-hydroxyvitamin D-1alpha-hydroxylase (1alpha-OHase) from 10 days of age. These phenotypes could not be explained by currently known regulators of mineral homeostasis, indicating that FGF23 is essential for normal phosphate and vitamin D metabolism.
Figures
References
-
- Brown EM, et al. Cloning and characterization of an extracellular Ca(2+)-sensing receptor from bovine parathyroid. Nature. 1993;366:575–580. - PubMed
-
- Fitzpatrick, L.A., and Bilezikian, J.P. 1996. Actions of parathyroid hormone. In Principles of bone biology. J.P. Bilezikian, L.G. Raisz, and G.A. Rodan, editors. Academic Press Inc. San Diego, California, USA. 339–346.
-
- Drezner MK. PHEX gene and hypophosphatemia. Kidney Int. 2000;57:9–18. - PubMed
-
- Econs MJ, Drezner MK. Tumor-induced osteomalacia—unveiling a new hormone. N. Engl. J. Med. 1994;330:1679–1681. - PubMed
-
- The ADHR Consortium. Autosomal dominant hypophosphataemic rickets is associated with mutations in FGF23. Nat. Genet. 2000;26:345–348. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
