

The calcium-sensing receptor regulates mammary gland parathyroid hormone-related protein production and calcium transport
- PMID: 14966569
- PMCID: PMC338258
- DOI: 10.1172/JCI18776
The calcium-sensing receptor regulates mammary gland parathyroid hormone-related protein production and calcium transport
Abstract
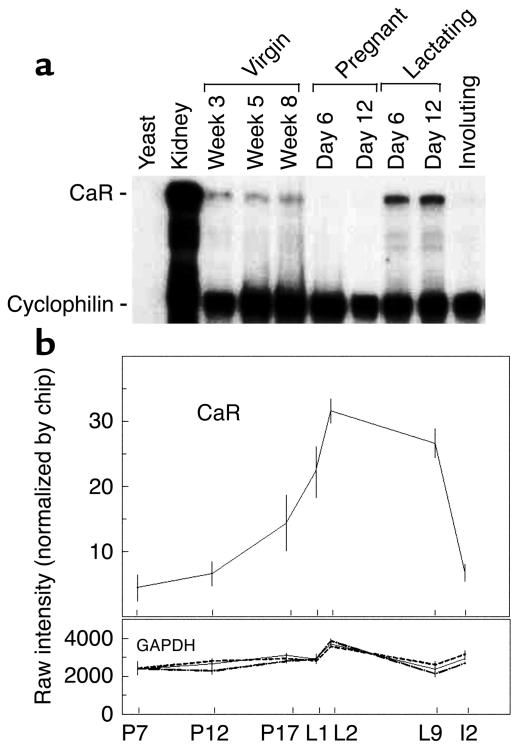
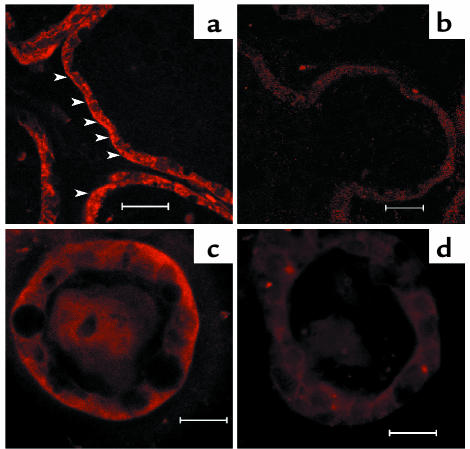
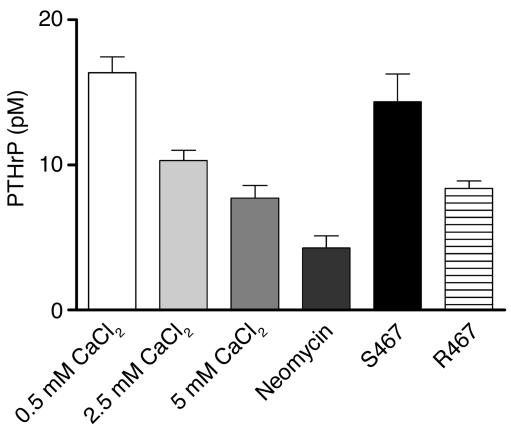
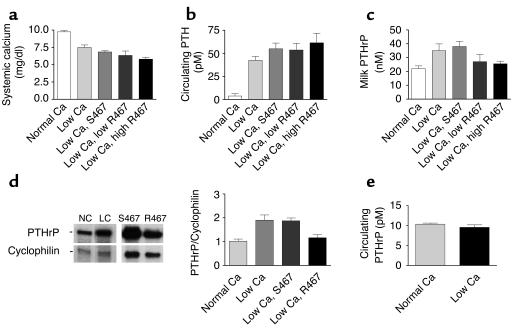
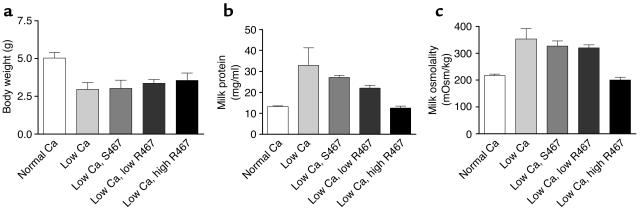
The transfer of calcium from mother to milk during lactation is poorly understood. In this report, we demonstrate that parathyroid hormone-related protein (PTHrP) production and calcium transport in mammary epithelial cells are regulated by extracellular calcium acting through the calcium-sensing receptor (CaR). The CaR becomes expressed on mammary epithelial cells at the transition from pregnancy to lactation. Increasing concentrations of calcium, neomycin, and a calcimimetic compound suppress PTHrP secretion by mammary epithelial cells in vitro, whereas in vivo, systemic hypocalcemia increases PTHrP production, an effect that can be prevented by treatment with a calcimimetic. Hypocalcemia also reduces overall milk production and calcium content, while increasing milk osmolality and protein concentrations. The changes in milk calcium content, milk osmolality, and milk protein concentration were mitigated by calcimimetic infusions. Finally, in a three-dimensional culture system that recapitulates the lactating alveolus, activation of the basolateral CaR increases transcellular calcium transport independent of its effect on PTHrP. We conclude that the lactating mammary gland can sense calcium and adjusts its secretion of calcium, PTHrP, and perhaps water in response to changes in extracellular calcium concentration. We believe this defines a homeostatic system that helps to match milk production to the availability of calcium.
Figures


References
-
- Brown EM, et al. Cloning and characterization of an extracellular Ca(2+)-sensing receptor from bovine parathyroid. Nature. 1993;366:575–580. - PubMed
-
- Brown EM, MacLeod RJ. Extracellular calcium sensing and extracellular calcium signaling. Physiol. Rev. 2001;81:239–297. - PubMed
-
- Chattopadhyay N, Mithal A, Brown EM. The calcium-sensing receptor: a window into the physiology and pathophysiology of mineral ion metabolism. Endocr. Rev. 1996;17:289–307. - PubMed
-
- Brown EM. Extracellular Ca2+ sensing, regulation of parathyroid cell function, and role of Ca2+ and other ions as extracellular (first) messengers. Physiol. Rev. 1991;71:371–411. - PubMed
-
- Conklin BR, Bourne HR. Homeostatic signals. Marriage of the flytrap and the serpent. Nature. 1994;367:22. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
