

Glutamate-183 in the conserved TGES motif of domain A of sarcoplasmic reticulum Ca2+-ATPase assists in catalysis of E2/E2P partial reactions
- PMID: 14970331
- PMCID: PMC365696
- DOI: 10.1073/pnas.0400122101
Glutamate-183 in the conserved TGES motif of domain A of sarcoplasmic reticulum Ca2+-ATPase assists in catalysis of E2/E2P partial reactions
Abstract
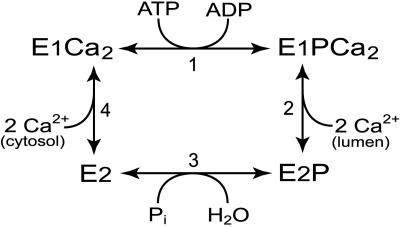
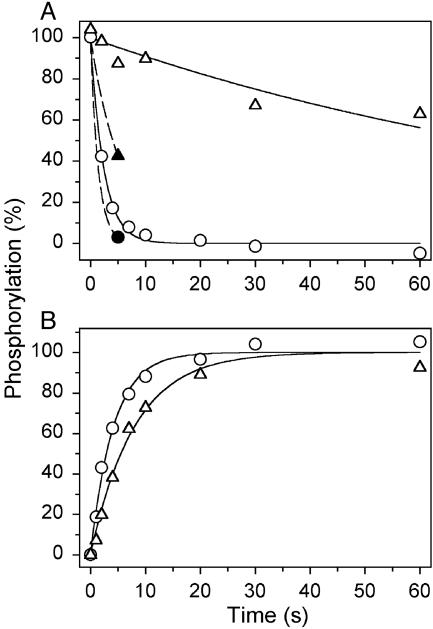
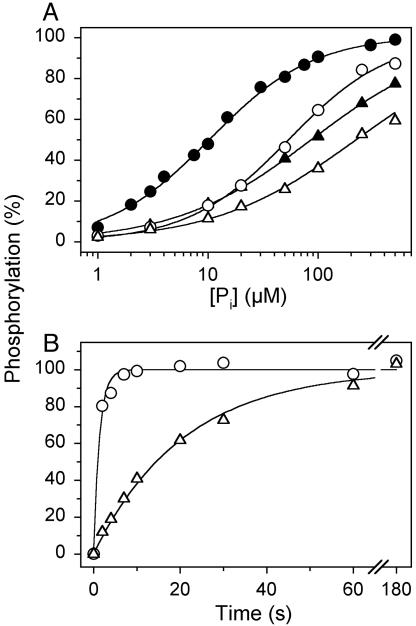
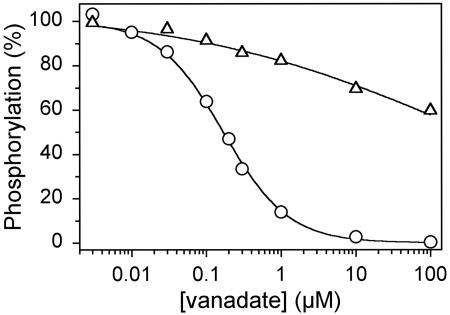
The recently determined crystal structures of the sarcoplasmic reticulum Ca(2+)-ATPase show that in the E(1)Ca(2) form, domain A is almost isolated from the other cytoplasmic domains, P and N, whereas in E(2), domain A has approached domains P and N, with E183 of the highly conserved P-type ATPase signature sequence TGES in domain A now being close to the phosphorylated aspartate in domain P, thus raising the question whether E183 acquires a catalytic role in E(2) and E(2)P conformations. This study compares the partial reactions of mutant E183A and wild-type Ca(2+)-ATPase, using transient and steady-state kinetic measurements. It is demonstrated that dephosphorylation of the E(2)P phosphoenzyme intermediate, as well as reverse phosphorylation of E(2) with P(i), is severely inhibited in the mutant. Furthermore, the apparent affinity of E(2) for the phosphoryl transition state analog vanadate is reduced by three orders of magnitude, consistent with a destabilization of the transition state complex, and the mutant displays reduced apparent affinity for P(i) in the E(2) form. The E(1)Ca(2) conformation, on the other hand, shows normal phosphorylation with ATP and normal Ca(2+) binding properties, and the rates of the conformational transitions E(1)PCa(2) --> E(2)P and E(2) --> E(1)Ca(2) are only 2- to 3-fold reduced, relative to wild type. These results, which likely can be generalized to other P-type ATPases, indicate that E183 is critical for the phosphatase function of E(2) and E(2)P, possibly interacting with the phosphoryl group or attacking water in the transition state complex, but is of little functional importance in E(1) and E(1)P.
Figures

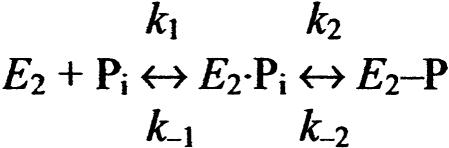

References
-
- de Meis, L. & Vianna, A. L. (1979) Annu. Rev. Biochem. 48, 275-292. - PubMed
-
- Inesi, G., Sumbilla, C. & Kirtley, M. E. (1990) Physiol. Rev. 70, 749-776. - PubMed
-
- MacLennan, D. H., Rice, W. J. & Green, N. M. (1997) J. Biol. Chem. 272, 28815-28818. - PubMed
-
- Kaplan, J. H. (2002) Annu. Rev. Biochem. 71, 511-535. - PubMed
-
- Morsomme, P., Slayman, C. W. & Goffeau, A. (2000) Biochim. Biophys. Acta 1469, 133-157. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
