

Identification of multivesicular bodies as prevacuolar compartments in Nicotiana tabacum BY-2 cells
- PMID: 14973159
- PMCID: PMC385280
- DOI: 10.1105/tpc.019703
Identification of multivesicular bodies as prevacuolar compartments in Nicotiana tabacum BY-2 cells
Abstract
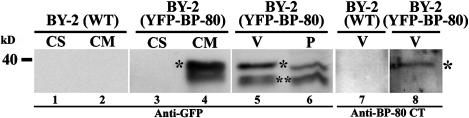
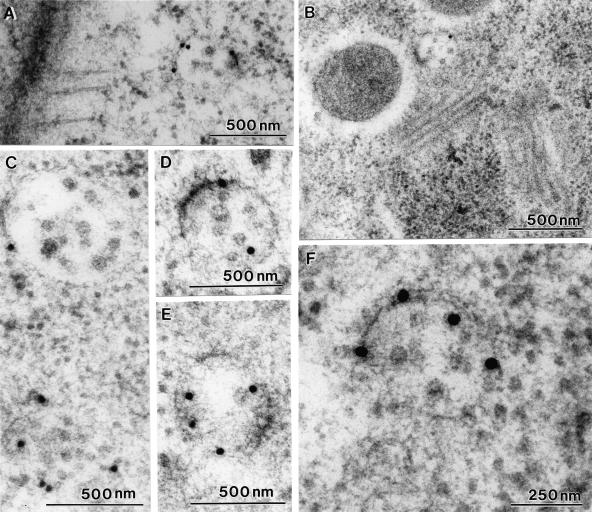
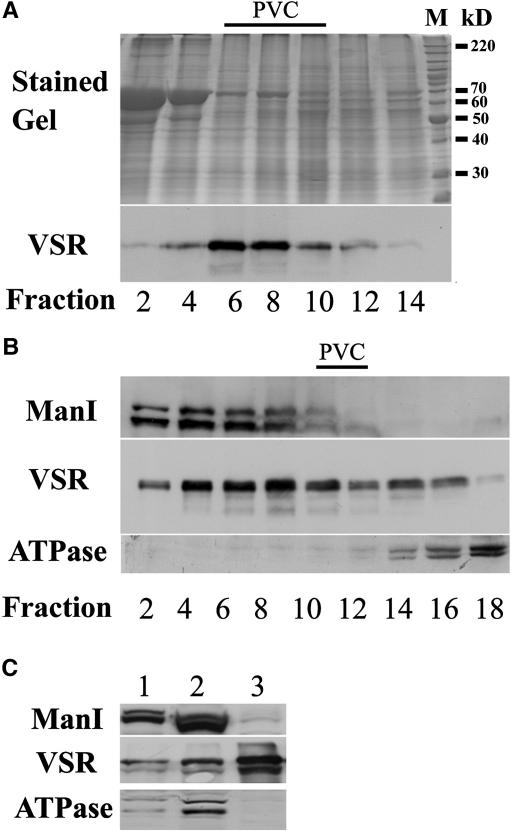
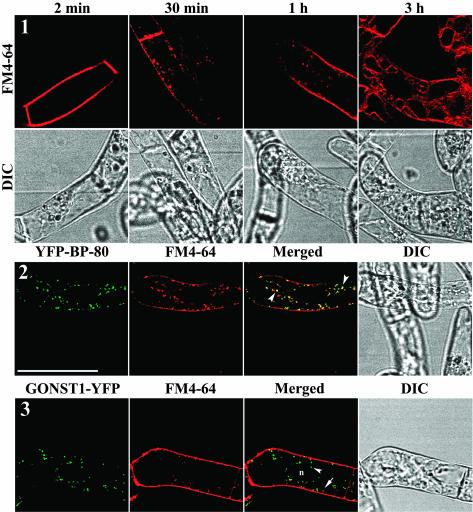
Little is known about the dynamics and molecular components of plant prevacuolar compartments (PVCs). We have demonstrated recently that vacuolar sorting receptor (VSR) proteins are concentrated on PVCs. In this study, we generated transgenic Nicotiana tabacum (tobacco) BY-2 cell lines expressing two yellow fluorescent protein (YFP)-fusion reporters that mark PVC and Golgi organelles. Both transgenic cell lines exhibited typical punctate YFP signals corresponding to distinct PVC and Golgi organelles because the PVC reporter colocalized with VSR proteins, whereas the Golgi marker colocalized with mannosidase I in confocal immunofluorescence. Brefeldin A induced the YFP-labeled Golgi stacks but not the YFP-marked PVCs to form typical enlarged structures. By contrast, wortmannin caused YFP-labeled PVCs but not YFP-labeled Golgi stacks to vacuolate. VSR antibodies labeled multivesicular bodies (MVBs) on thin sections prepared from high-pressure frozen/freeze substituted samples, and the enlarged PVCs also were indentified as MVBs. MVBs were further purified from BY-2 cells and found to contain VSR proteins via immunogold negative staining. Similar to YFP-labeled Golgi stacks, YFP-labeled PVCs are mobile organelles in BY-2 cells. Thus, we have unequivocally identified MVBs as PVCs in N. tabacum BY-2 cells. Uptake studies with the styryl dye FM4-64 strongly indicate that PVCs also lie on the endocytic pathway of BY-2 cells.
Figures










References
-
- Ahmed, S.U., Rojo, E., Kovaleva, V., Venkataraman, S., Dombrowski, J.E., Matsuoka, K., and Raikhel, N.V. (2000). The plant vacuolar sorting receptor AtELP is involved in transport of NH(2)-terminal propeptide-containing vacuolar proteins in Arabidopsis thaliana. J. Cell Biol. 149, 1335–1344. - PMC - PubMed
-
- Aniento, F., Helms, J.B., and Memon, A.R. (2003). How to make a vesicle: Coat protein-membrane interactions. Annu. Plant Rev. 9, 36–62.
-
- Barr, F.A. (2002). Inheritance of the endoplasmic reticulum and Golgi apparatus. Curr. Opin. Cell Biol. 14, 496–499. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
