

Intact type 1 immunity and immune-associated coagulative responses in mice lacking IFN gamma-inducible fibrinogen-like protein 2
- PMID: 14976252
- PMCID: PMC365735
- DOI: 10.1073/pnas.0308369101
Intact type 1 immunity and immune-associated coagulative responses in mice lacking IFN gamma-inducible fibrinogen-like protein 2
Abstract
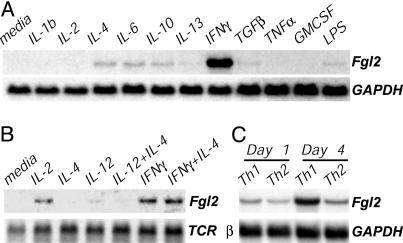
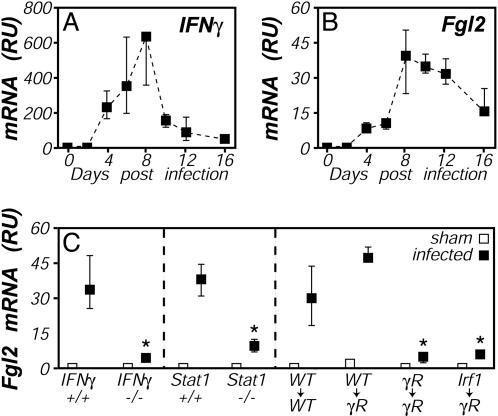
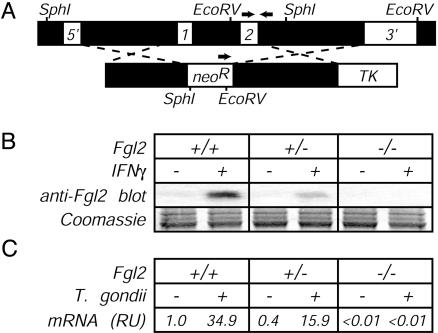
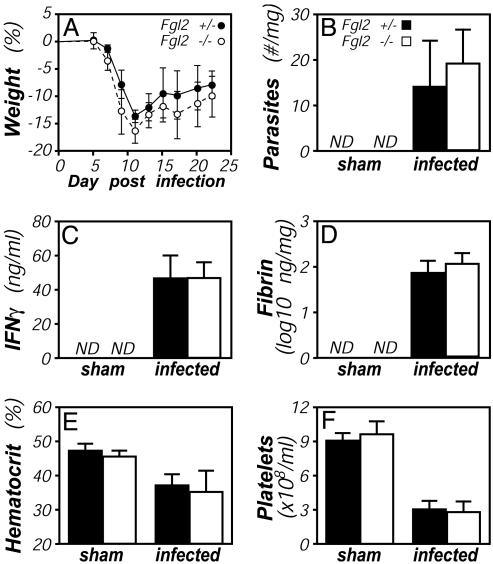
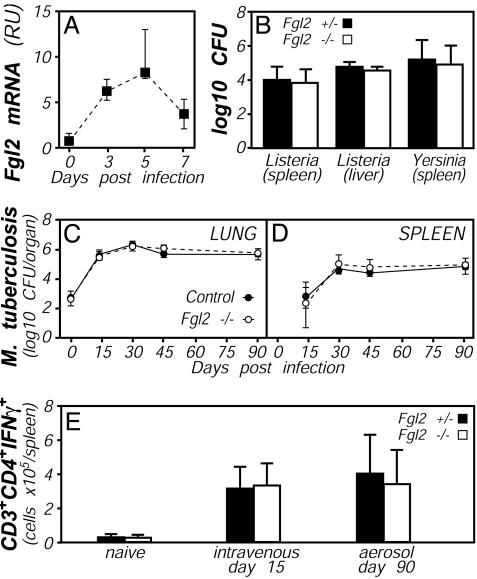
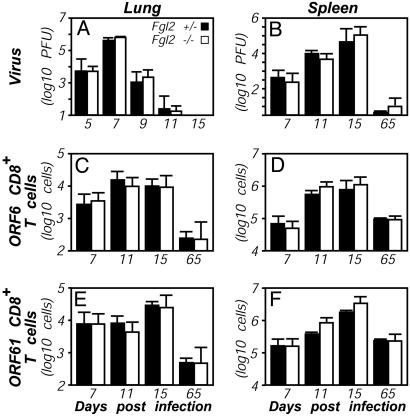
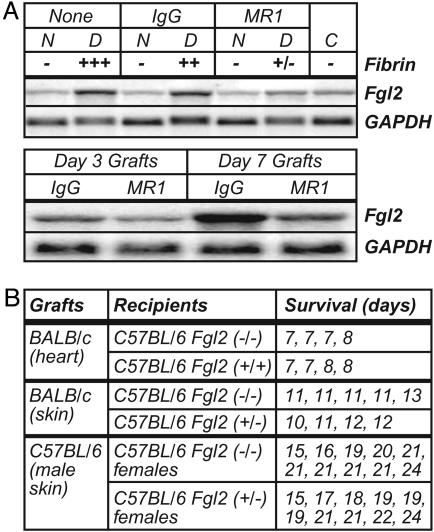
Fibrinogen-like protein 2 (Fgl2, fibroleukin) is a leukocyte product that exhibits significant homology to secreted proteins of diverse function, including growth factors, lectins, and components of extracellular matrix. Prior studies found that Fgl2 is IFN gamma-inducible, possesses direct coagulant activity, and inhibits T cell proliferation and dendritic cell maturation in vitro. Here, we demonstrate that Fgl2 expression is up-regulated during type 1 immunity in vivo and establish that such up-regulation is IFN gamma-, signal transducer and activation of transcription protein 1-, and IFN response factor 1-dependent. To investigate functional roles for Fgl2 during type 1 immunity, we generated Fgl2-deficient mice. Those animals are born at predicted Mendelian frequencies, appear overtly healthy, and contain normal numbers and frequencies of lymphoid cells. Although Fgl2 is IFN gamma-inducible and putatively regulates T cell activation/proliferation, we demonstrate that Fgl2-deficient and control mice exhibit similar degrees of T cell expansion, immunopathology, and/or pathogen burdens during protozoan (Toxoplasma gondii), bacterial (Yersinia enterocolitica, Listeria monocytogenes, and Mycobacterium tuberculosis), and viral (murine gamma-herpesvirus-68 and Sendai) infections. Fgl2-deficient mice also reject allografts with similar kinetics as control mice. Moreover, despite prior reports that Fgl2 functions as a procoagulant enzyme, we demonstrate that Fgl2-deficient and control mice produce similar levels of fibrin, a product of the coagulation cascade, during T. gondii infection and allograft rejection. Together, our findings suggest that Fgl2, although highly conserved and IFN gamma-inducible, is not a critical mediator of either type 1 immunity or immune-associated coagulant activity.
Figures
References
-
- Ruegg, C. & Pytela, R. (1995) Gene 160, 257-262. - PubMed
-
- Marazzi, S., Blum, S., Hartmann, R., Gundersen, D., Schreyer, M., Argraves, S., von Fliedner, V., Pytela, R. & Ruegg, C. (1998) J. Immunol. 161, 138-147. - PubMed
-
- Lafuse, W. P., Castle, L., Brown, D. & Zwilling, B. S. (1995) Cell. Immunol. 163, 187-190. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
