

Role of MyD88 in diminished tumor necrosis factor alpha production by newborn mononuclear cells in response to lipopolysaccharide
- PMID: 14977922
- PMCID: PMC355999
- DOI: 10.1128/IAI.72.3.1223-1229.2004
Role of MyD88 in diminished tumor necrosis factor alpha production by newborn mononuclear cells in response to lipopolysaccharide
Abstract
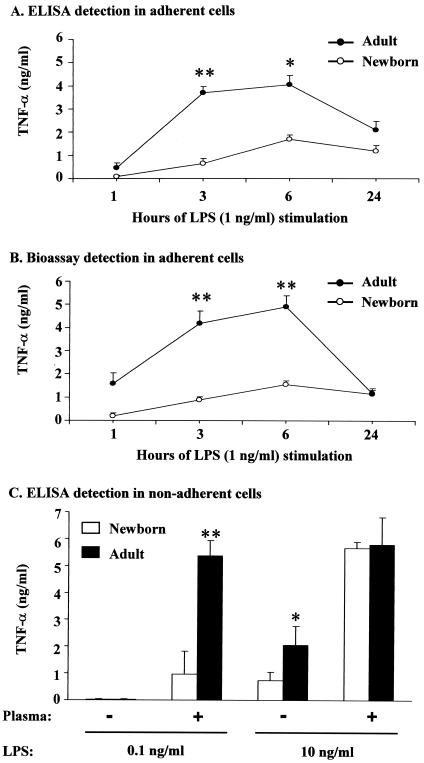
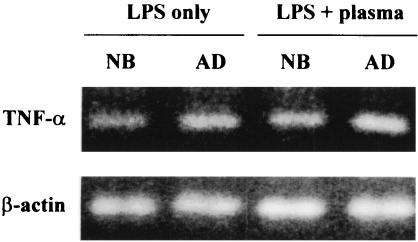
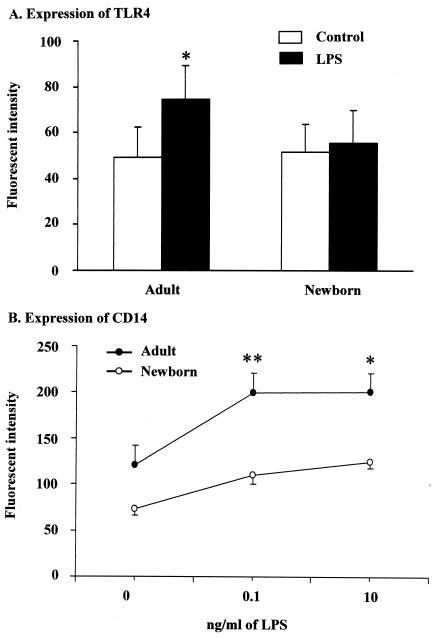
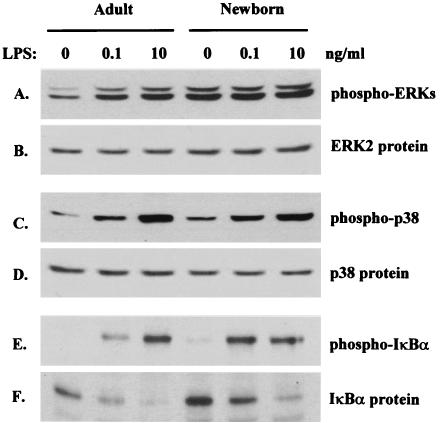
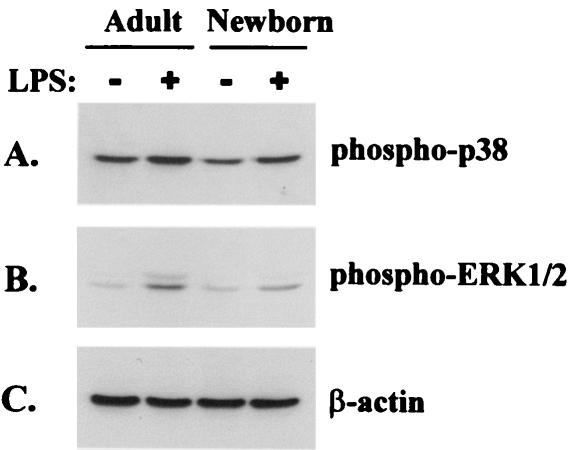
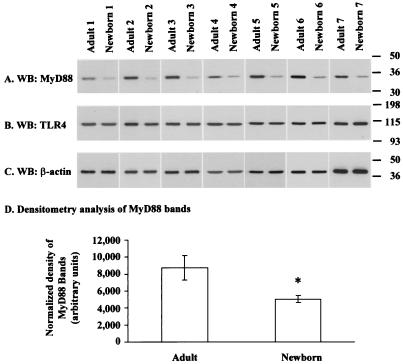
Human newborns are more susceptible than adults to infection by gram-negative bacteria. We hypothesized that this susceptibility may be associated with a decreased response by leukocytes to lipopolysaccharide (LPS). In this study, we compared LPS-induced secretion of tumor necrosis factor alpha (TNF-alpha) by mononuclear cells (MNC) from adult peripheral blood and newborn umbilical cord blood in vitro and attempted to determine the mechanisms involved in its regulation. At a high concentration of LPS (10 ng/ml) and in the presence of autologous plasma, MNC from adults and newborns secreted similar amounts of TNF-alpha. However, in the absence of plasma, MNC from newborns secreted significantly less TNF-alpha compared to MNC from adults. Moreover, at a low concentration of LPS (0.1 ng/ml) and in the presence of plasma, TNF-alpha secretion was significantly lower for newborn MNC compared to adult MNC. Adults and newborns had similar numbers of CD14 and Toll-like receptor 4 (TLR-4)-positive cells as measured by flow cytometry. However, the intensity of the CD14 marker was greater for adult than for newborn cells. Incubation of cells with LPS led to an increase in CD14 and TLR-4 intensity for adult cells but not for newborn cells. The effect of LPS stimulation of adult or newborn cells was similar for ERK, p38, and IkappaBalpha phosphorylation, as well as IkappaBalpha degradation. Finally, we assessed levels of the TLR-4 adapter protein, the myeloid differentiation antigen 88 (MyD88). We found a direct relation between adult and newborn TNF-alpha secretion and MyD88, which was significantly decreased in newborn monocytes. Since TLR-4 signals intracellularly through the adapter protein, MyD88, we hypothesize that MyD88-dependent factors are responsible for delayed and decreased TNF-alpha secretion in newborn monocytes.
Figures
References
-
- Aida, Y., and M. J. Pabst. 1990. Priming of neutrophils by lipopolysaccharide for enhanced release of superoxide: requirement for plasma but not for tumor necrosis factor-α. J. Immunol. 145:3017-3025. - PubMed
-
- Alexopoulou, L., A. C. Holt, R. Medzhitov, and R. A. Flavell. 2001. Recognition of double-stranded RNA and activation of NF-κB by Toll-like receptor 3. Nature 413:732-738. - PubMed
-
- Beutler, B. 2000. Tlr4: central component of the sole mammalian LPS sensor. Curr. Opin. Immunol. 12:20-26. - PubMed
-
- Beutler, B. 1990. TNF in pathophysiology: biosynthetic regulation. J. Investig. Dermatol. 95:81S-84S. - PubMed
-
- Bohmer, R. H., L. S. Trinkle, and J. L. Staneck. 1992. Dose effects of LPS on neutrophils in a whole blood flow cytometric assay of phagocytosis and oxidative burst. Cytometry 13:525-531. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
