

Serum leptin level is a regulator of bone mass
- PMID: 14978271
- PMCID: PMC365777
- DOI: 10.1073/pnas.0308744101
Serum leptin level is a regulator of bone mass
Abstract
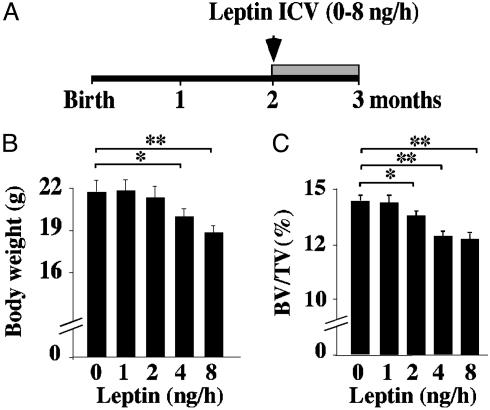
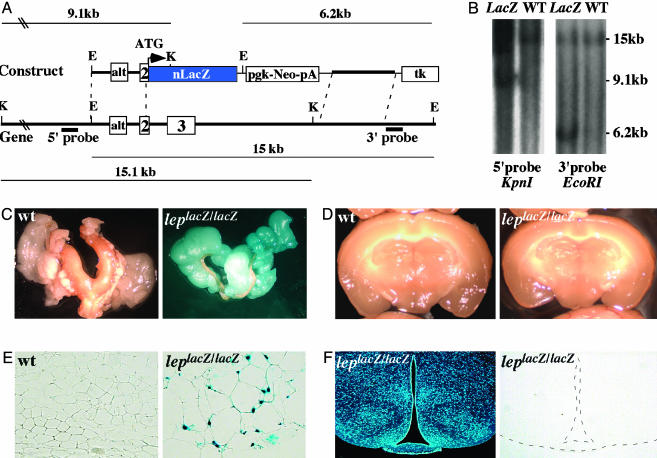
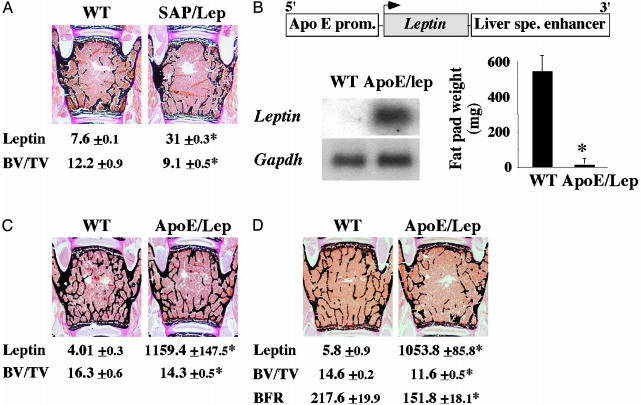
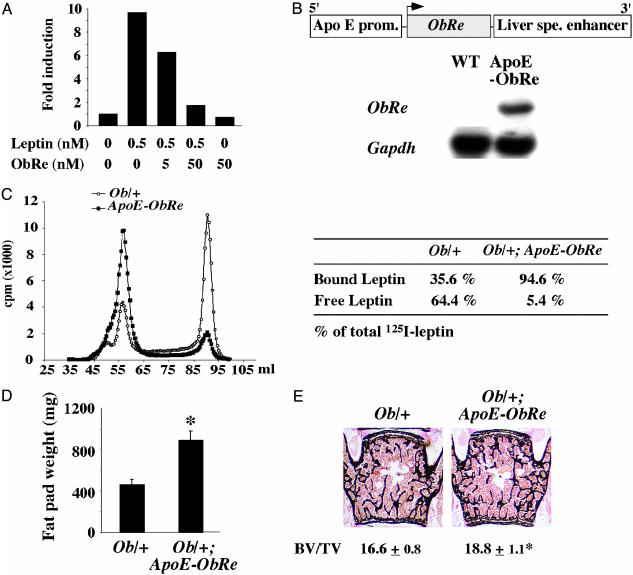
Leptin is a powerful inhibitor of bone formation in vivo. This antiosteogenic function involves leptin binding to its receptors on ventromedial hypothalamic neurons, the autonomous nervous system and beta-adrenergic receptors on osteoblasts. However, the mechanisms whereby leptin controls the function of ventromedial hypothalamic antiosteogenic neurons remain unclear. In this study, we compared the ability of leptin to regulate body weight and bone mass and show that leptin antiosteogenic and anorexigenic functions are affected by similar amounts of leptin. Using a knock-in of LacZ in the leptin locus, we failed to detect any leptin synthesis in the central nervous system. However, increasing serum leptin level, even dramatically, reduced bone mass. Conversely, reducing serum-free leptin level by overexpressing a soluble receptor for leptin increased bone mass. Congruent with these results, the high bone mass of lipodystrophic mice could be corrected by restoring serum leptin level, suggesting that leptin is an adipocyte product both necessary and sufficient to control bone mass. Consistent with the high bone mass phenotype of lipodystrophic mice, we observed an advanced bone age, an indirect reflection of premature bone formation, in lipodystrophic patients. Taken together, these results indicate that adipocyte-derived circulating leptin is a determinant of bone formation and suggests that leptin antiosteogenic function is conserved in vertebrates.
Figures
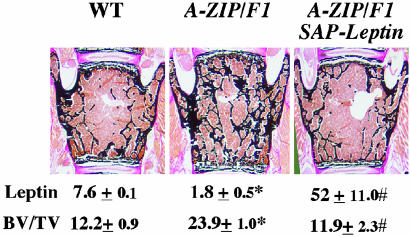
References
-
- Ducy, P., Amling, M., Takeda, S., Priemel, M., Schilling, A. F., Beil, T., Shen, J., Vinson, C., Rueger, J. M. & Karsenty, G. (2000) Cell 100, 197-207. - PubMed
-
- Takeda, S., Elefteriou, F., Levasseur, R., Liu, X., Zhao, L., Parker, K. L., Armstrong, D., Ducy, P. & Karsenty, G. (2002) Cell 111, 305-317. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
